


Abstract
Little information is available about the effects of exposure to radiofrequency electromagnetic fields (RF) on cerebral microcirculation during rat developmental stages. We investigated whether the permeability of the blood–brain barrier (BBB) in juvenile and young adult rats was modified during local cortex exposure to RF under non-thermal conditions. The cortex tissue targeted was locally exposed to 1457 MHz RF at an average specific absorption rate of 2.0 W/kg in the target area for 50 min and permeability changes in the BBB of the pia mater were measured directly, using intravital fluorescence microscopy. There was no significant difference in extravasation of intravenously-injected dye between exposed and sham-exposed groups of either category of rats. No histological evidence of albumin leakage was found in any of the brains just after exposure, indicating that no traces of BBB disruption remained. These findings suggest that no dynamic changes occurred in BBB permeability of the rats at either of these developmental stages, even during local RF exposure at non-thermal levels.
- Radiofrequency electromagnetic field
- local exposure
- blood-brain barrier
- temperature
- juvenile
- young adult
- development stage
Recently, there has been some concern regarding the possible adverse effects of radiofrequency electromagnetic fields (RF) on human health. In particular, the impact of RF exposure on the health of children during their development is an important issue due to the recent widespread use of smartphones among the younger generation. Another reason may be anxiety about age-related differences in sensitivity to chemical/physical factors. Many experimental studies in animals suggest higher sensitivity to these factors during the developmental stage than in adults. For example, ethanol affects synaptic activity in developing rat hippocampus with higher sensitivity than in adults (1, 2). Among physical factors, rat brains are known to have age-dependent sensitivity to cellular injury caused by irradiation (3). Therefore, RF is considered to be a physical factor likely to impact health in developing brains.
Further studies are required to evaluate the biological effects of RF on developing brains (4, 5). To provide protection against known adverse health effects, the International Commission on Non-Ionizing Radiation Protection (ICNIRP) established guidelines specifying the basic restriction values in terms of specific absorption rates (SAR, W/kg) for tissue or the whole body (5). For example, the restriction values for general public exposure concerning local and whole-body RF exposure (100 kHz-10 GHz) are 2 and 0.08 W/kg, respectively. These values include a safety factor to allow for the difference between developing children and adults. However, these figures are based on evidence of RF-induced thermal effects and dosimetric readings of thermal changes (6-9). Few biological effects, such as neuro- and vessel toxicities in the developmental stage, are reflected in the guidelines.
Several research groups have evaluated the biological effects of RF exposure in rodents during the developmental stage (10-14). Paulraj and Behari found an increase in brain glial cell populations of rats in the developmental stage (35 to 70 days old) following chronic whole-body RF exposure at 0.11 W/kg (10). Kumlin et al. also reported that whole-body RF exposure of developing rats (for five weeks) improved their learning and memory in the water maze test (11). However, in contrast, Klose et al. found that daily RF exposure to the head region of rats had no harmful effects on their learning skill or behavior, even in juveniles (12). Kuribayashi et al. also failed to find any adverse effects on rats during the developmental stage. Local brain exposure to RF did not affect the blood–brain barrier (BBB) of immature and young rats (13). Therefore, the issue of the effects of RF during the developmental stage is still controversial. In particular, there is little information about the possibility of dynamic variations during RF exposure, likely to appear as transient phenomena only.
The aim of the present study was to investigate whether dynamic variations in BBB permeability were observed in juvenile and young adult rat brains during local cortex exposure to RF. Our group has already examined changes in BBB permeability in real-time during RF exposure of adult rats, but not for rats in the developmental stage. We used intravital fluorescence microscopy developed for rats (15) and a figure-8 loop antenna to provide the local cortex exposure (16).
Materials and Methods
Animals. Male Sprague-Dawley rats of two different ages (n=31, 4-week-old-juvenile rats, 127±4 g; n=28, 8-week-old young adult rats, 335±4 g; Japan SLC, Inc., Shizuoka, Japan) were used for this experiment. They were fed a standard pellet diet, given water ad libitum, and housed in an animal room with a 12-h light/dark cycle, at a temperature of 23.0±1°C and a relative humidity of 50±10%. All experimental procedures were conducted in accordance with the ethical guidelines for animal experiments at the National Institute of Public Health, Japan.
Preparation of closed cranial window. The closed cranial window setup (CCW) was implanted into the parietal region of each rat at least one week before the observation began (Figure 1A). The CCW was developed to observe pial microcirculation under RF exposure conditions (15, 17, 18). In brief, the rats were anesthetized with an intramuscular injection of ketamine (100 mg/kg) and xylazine (10 mg/kg). After removal of the skin, connective tissues, and a 7.5-mm circular patch of skull from the parietal region, the CCW (circular cover-glass 7.0 mm in diameter) was inserted into the hole in the skull and fixed.
Definition of target area and local RF exposure. The target cortex tissue was locally exposed to 1439 MHz RF (signal type: Personal Digital Cellular signal) at several averaged SAR for the target cortex using a figure-8 loop antenna positioned 5 mm above the CCW (16).
The target area was defined as a discoid area of rat parietal cortex tissue, located just under the CCW, with its center 1.5 mm posterior to the bregma and 1.5 mm lateral (left) fthe midline. The area included the pia mater (7.5 mm diameter, 0.5 mm depth; Figure 1A) in which we evaluated physiological responses during local RF exposure.
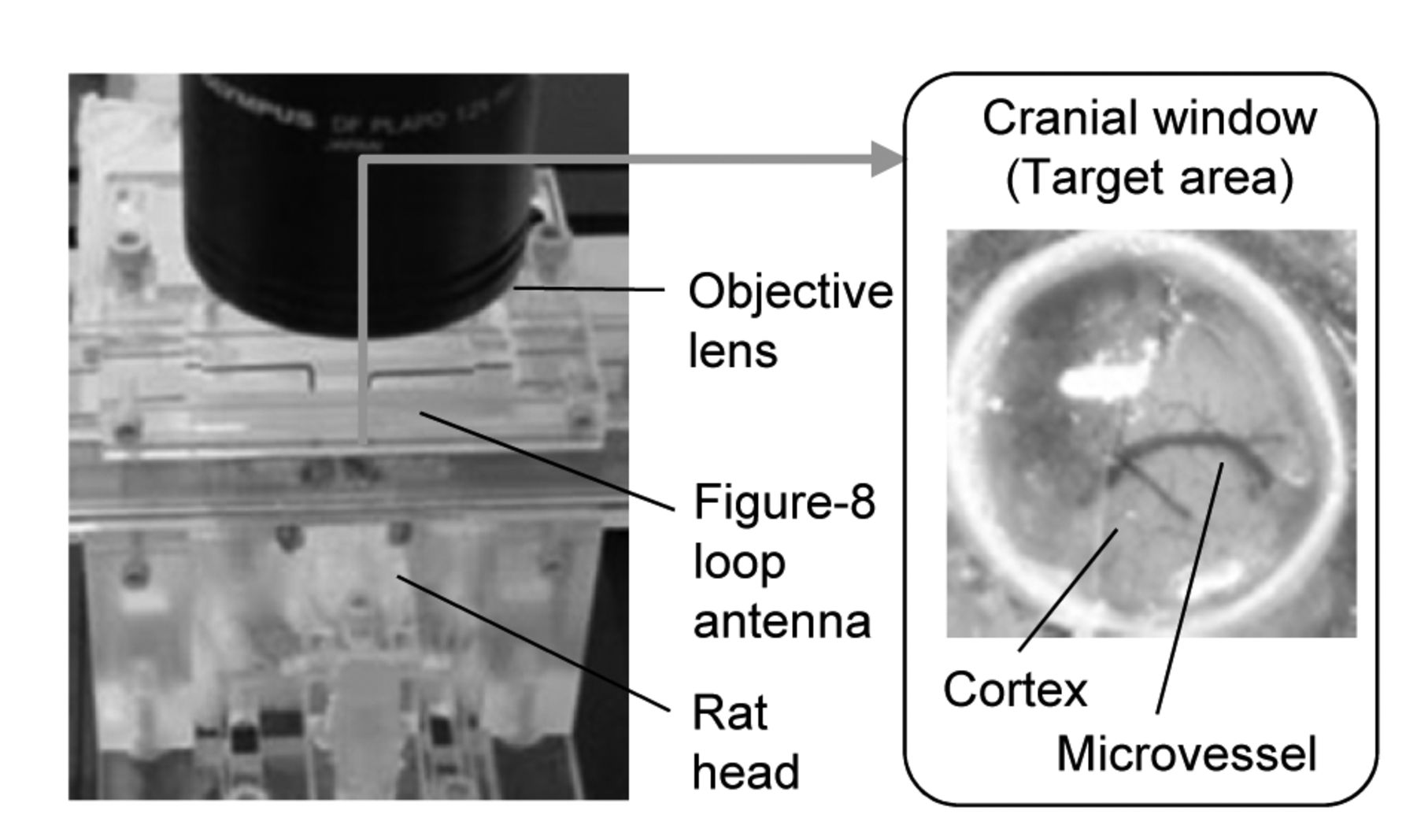
Real-time observation in vivo. The closed cranial window was implanted in the rat parietal region at least one week before the experiment. The target area was identified as a circular area of parietal cortex tissue located just under the window (7.5 mm diameter, 0.5 mm depth). The antenna was placed exactly over the parietal region using a manipulator. Rat pial microcirculation was observed through holes in the antenna in real-time using intravital microscopy.
RF exposure intensities were estimated and set at 2.0, 6.8, 25.4, 84.6, 254 W/kg SAR in the target area (TASAR) for juvenile rats, and 2.0, 7.4, 28.0, 92.9, 278 W/kg TASAR for young adult rats. For juvenile rats, the relative values of brain- and whole-body-averaged SARs for the TASAR of 2.0 W/kg were 0.59 and 0.057 W/kg, respectively. For young adult rats, these averaged SARs were 0.54 and 0.03 W/kg, respectively.
The exposure was performed for 50 min at 2.0 W/kg to record physiological parameters and for 10 min at the other SARs in turn to obtain a relationship between TASAR and temperature changes in the target area and rectum (Figure 1B). Sham-exposed rats were also prepared using the same system, but without RF exposure (0 W/kg).
Real-time measurement of physiological parameters. Temperatures in two regions (target area and rectum) and BBB permeability were measured before, during, and just after RF exposure, using our previous method (19). In brief, the rats were anesthetized with an intramuscular injection of ketamine (100 mg/kg) and xylazine (10 mg/kg) and a subcutaneous injection of pentobarbital (12.5 mg/kg). The rat was placed on a heating pad circulating warmed water (42°C).
Temperatures in the target area and rectum were measured using optical thermometer probes (m600; Lumasense Technologies, Santa Clara, CA, USA) placed on the target and in the rectum. Temperatures were recorded throughout the experiment for 50 min exposure at 2.0 W/kg TASAR in sham and RF groups (n=3 animals each), and then measured during 10 min-RF exposures at the different TASARs mentioned above (n=2–6 animals each).
BBB permeability was evaluated as changes in the intensity of extravasated fluorescence dye, as in previous studies (17, 18, 20). In brief, after intravenous injection of fluorescein isothiocyanate (FITC)-dextran (70 kDa, 25 mg/kg; Sigma-Aldrich Co., St. Louis, MO, USA), the image of the pia mater within the target area was recorded through the intravital fluorescence microscopic system built during our previous study (18). An arbitrary area of the pia mater was chosen as the region of interest (ROI; 0.19 mm2, three areas per animal). The average fluorescence intensity of the ROI was measured offline every 10 min.
Histological examination. Rat brain exposed to RF was histologically examined as in previous studies (21). Briefly, immediately after the last measurements of microcirculatory parameters, the rat was transcardially perfused with 4% paraformaldehyde in phosphate-buffered saline. The brain was removed from the skull and its vertical cryosections (10 μm-thick) were taken from the frontal (between bregma −0.8 mm and −1.2 mm) and middle (between bregma −2.0 mm and −5.0 mm) brains. Immunohistochemistry of extravasated serum albumin was detected by immunohistochemistry with rabbit polyclonal antibody to human albumin (1:100 dilution, A0001; Dako, Japan Inc., Tokyo, Japan), which is known to react to rat albumin too (21). The presence of antibodies was detected using 3,3’-diaminobenzidine chromogen reaction (K3466; Dako), and nuclei were identified using hematoxylin.
Data analysis. A two-way repeated ANOVA and a Mann–Whitney U-test were used for statistical analysis to evaluate the differences between the sham- and RF-exposed groups. A value of p<0.05 was considered statistically significant.
Results
Temperature conditions. Temperatures in the target area and rectum were measured during exposure to verify that the findings of this experiment were not distorted by thermal conditions caused by RF exposure at 2.0 W/kg TASAR.
Basal temperatures in both regions and rats of both ages at the beginning of exposure were stabilized, without any significant difference between sham and RF groups (n=3 animals per group). In juvenile rats, the average temperatures in the target area in sham- and RF-treated groups were 34.6±1.1°C and 35.1±0.8°C, respectively. The average rectal temperatures for sham- and RF-treated groups were 35.4±0.3°C and 36.1±0.7°C, respectively. Similar results were obtained in young adult rats. The average temperatures in the target area in sham- and RF-treated groups were 34.7±0.3°C and 35.0±0.1°C, respectively. The average rectal temperatures for sham- and RF-treated groups were 36.2±0.3°C and 36.2±0.1°C, respectively. There were no significant differences in either basal temperatures between the two exposed groups.
During RF exposure at 2.0 W/kg TASAR, there were no significant differences in target or rectal temperatures between sham-treated and RF-exposed groups in rats of either age group (Figure 2A). Temperatures in the target area and rectum in both groups changed slightly during 50 min exposure. However, the absolute values of these temperature changes were only within ±0.3°C and showed no significant difference between the two groups during the exposure time-course. These findings confirmed that local RF exposure at 2.0 W/kg did not cause any significant increase in temperature.
There was a clear relationship between the TASAR and temperature changes in the target area and rectum, but only in the RF-treated group (Figure 2B). In rats of both age groups, the temperature in the target area increased at around 20 W/kg TASAR, whereas the rectal temperature only increased around 100 W/kg TASAR or more. These TASARs were at least 10-times higher than the value used to evaluate biological effects.
Changes in BBB permeability. To investigate whether the BBB permeability was modified under local RF exposure at 2.0 W/kg TASAR, variations in fluorescence intensity in the target area were measured during exposure following intravenous injection of the fluorescein isothiocyanate (FITC)-dextran fluorescence dye (70 kDa).
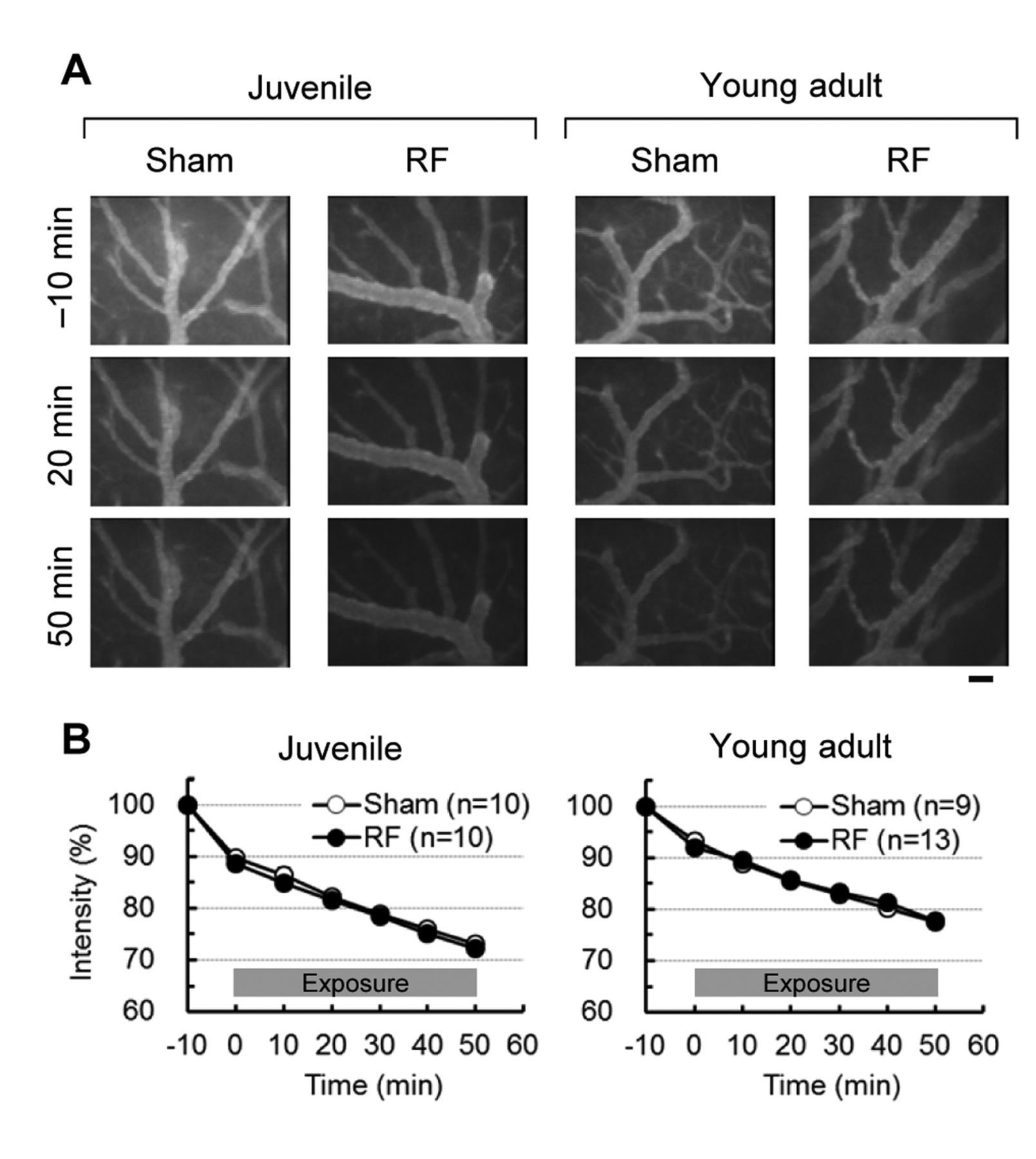
We have previously reported that extravasation of fluorescent dye was observed under intravital microscopy used for this experiment in a positive control using mannitol treatment (15, 18). Diffusion of the dye into the parenchyma around the microvessels resulted in accumulation of dye in that area. This accumulation was detected as an increase in fluorescence intensity in the target area. However, no extravasated fluorescent dye was observed in any regions of the target area in either sham- or RF-treated groups (Figure 3A). In juvenile rats, fluorescence intensity simply decreased time-dependently during the observation period (Figure 3B), which was attributed to dye clearance from the plasma (17). In addition, there was no significant difference in fluorescence intensity between the two groups. Similar results were obtained in young adult rats (Figure 3B). No significant difference in intensity changes was found between the two groups. These results revealed that no change in BBB permeability to FITC-dextran was caused by local RF exposure at 2.0 W/kg TASAR.
Histological evaluation. If BBB permeability was modified during RF exposure at 2.0 W/kg TASAR, some traces caused by this modification should be found in the brain immediately after exposure. Traces of changes in BBB permeability were immunohistochemically-assessed by measuring the extravasation of serum albumin from cerebral vessels (Figure 4). No immunostained sites were found in any of the brain sections of juvenile or young adult animals in the sham- or RF-treated groups (three regions in three animals per group). The results showed that no albumin entered the parenchyma through the cerebral microvessels, even after RF exposure at 2.0 W/kg.
Discussion
This in vivo study using real-time observation in juvenile and young adult rats produced two main findings. Firstly, local RF exposure of a target cortex area to 2.0 W/kg TASAR did not induce any temperature increase in the target throughout the experiment in rats of either age group. Secondly, no extravasation of intravenously injected fluorescent dye was detected in any target cortex during exposure.
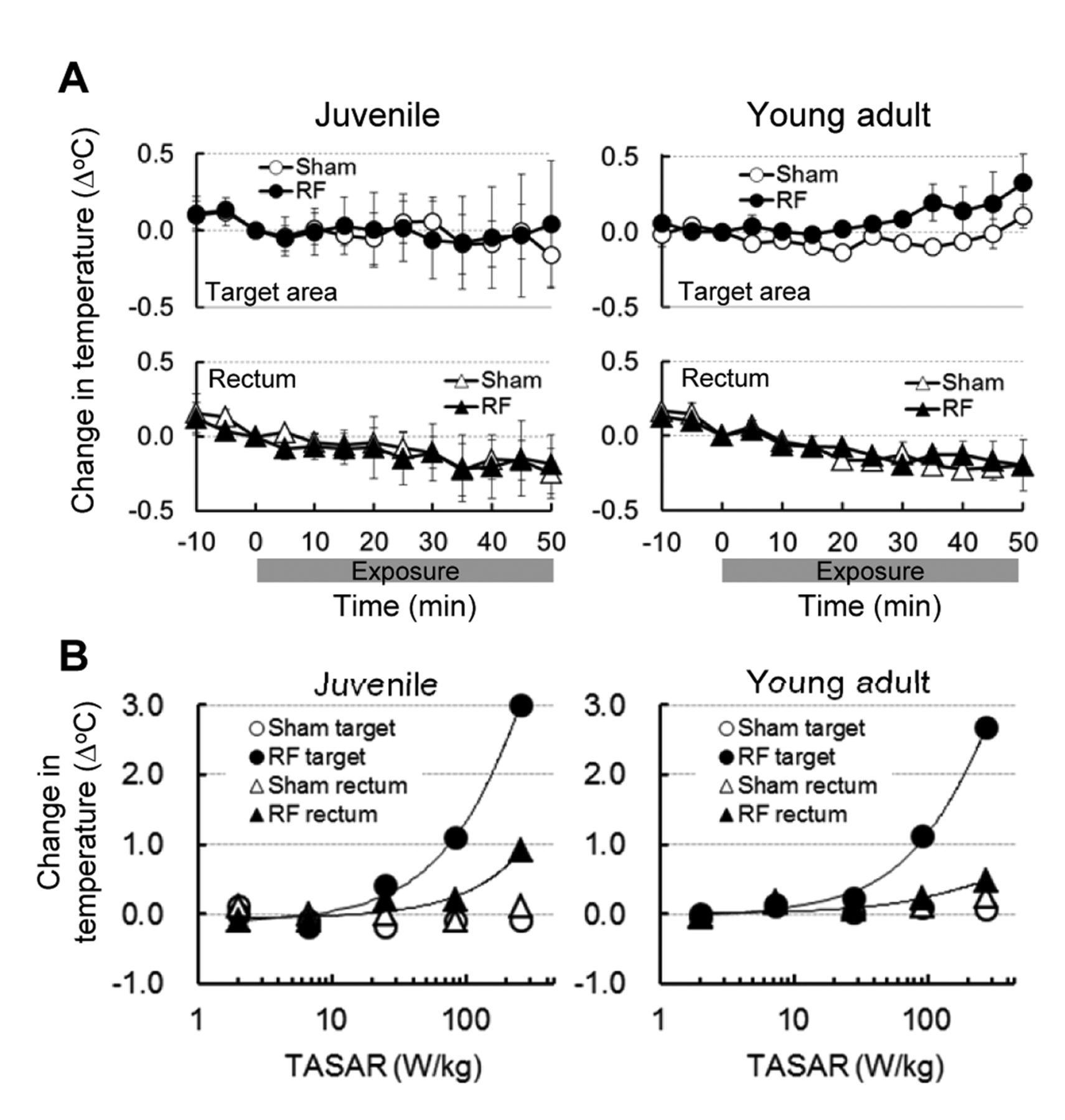
Temperature changes in the target area and rectum during radiofrequency electromagnetic field (RF) exposure. A: Temperature variations during 50 min-RF exposure at 2.0 W/kg specific absorption rate in the target area (TASAR). No significant difference in target or rectal temperatures between sham- and RF-exposed groups was found in 4-week-old juvenile rats nor 8-week-old young adult rats (n=3 animals per group). Values are the mean±SEM. B: Relationship between TASAR and temperature change. The value shows temperature changes during 10-min RF exposure at several TASARs. Values are the mean (n=2 to 6 animals per group).
This experiment focused on two key points for evaluating the biological effects of RF exposure. One was localizing the RF exposure to the cerebral tissue of young rats. The other was eliminating temperature elevation during local RF exposure, as we focused on the non-thermal biological effects of exposure. The figure-8 loop antenna we developed providing for effective local exposure of the rat parietal cortex without temperature elevation. The exposure intensities in the target cortex areas were estimated to be 35-fold for 4-week-old juvenile rats and 63-fold for 8-week-old young adult rats compared to whole-body exposure in our dosimetry results (16). This larger disparity in exposure intensity reflected the very low whole-body exposure even in juvenile rats. In fact, this disparity was confirmed in the relationship between the temperatures measured in the target area and rectum. At least 200 W/kg TASAR were required to increase the rectal temperature by 0.5°C, but only 40 W/kg TASAR in the target area. In addition, little heat generation was induced in the rectum via rat whole-body irradiaton except for the head, using similar exposure systems (19), suggesting effective local exposure of the young rat brains. Thanks to the local exposure method, the exposure at 2.0 W/kg TASAR certainly did not induce any temperature increase in the target area or rectum during the 50-min experiment. Therefore, it seems likely that the physiological responses in the study were obtained with little direct effect on whole-body exposure under non-thermal conditions.

Real-time observation of dye extravasation in target cortex tissue. A: Representative fluorescence images of the target area before, during, and after Radiofrequency electromagnetic field (RF) exposure. Fluorescein isothiocyanate dextran (70 kDa) was intravenously injected into the rats 25 min before RF exposure. Images were recorded in real time under an intravital fluorescence microscope. No extravasation of the dye was observed in 4-week-old juvenile nor in 8-week-old young adult rats. Bar: 50 μm. B: Time-course of fluorescence intensity in the target area. Intensity was monitored and averaged in regions of interest (three per animal) in the target area. No increase in average intensity was measured in juvenile nor in young adult rats. No significant differences were found between the sham- and RF-exposed groups (n=9 to 13 animals per group). Values are the mean±SEM.
This experiment did not detect any indications of permeability changes in the BBB during RF exposure at 2.0 W/kg TASAR. In particular, we evaluated the possibility of dynamic variations, likely to appear as transient phenomena during RF exposure in vivo. However, no extravasation of intravenously-injected fluorescence dye was directly observed during the 50-min RF exposure in target areas of rats in either age group. In addition, it was suggested that no dynamic changes in BBB permeability occurred in the whole brain, as no trace of extravasation of endogenous albumin (66 kDa), with a molecular size similar to that of fluorescein isothiocyanate (FITC)-dextran (70 kDa), was found in any regions of the cerebral slices, or in the target areas, even in juvenile rats. In our previous study on BBB permeability in 14-week-old adult rats, using the same methodology, no dynamic leakage of intravenously injected FITC-dextran was found during 50-min RF exposure at 2.0 W/kg TASAR without any temperature elevation (18). Therefore, these findings suggested that RF exposure at intensities below 2.0 W/kg TASAR did not affect BBB permeability in developing rats. Consequently, there was no difference between juvenile and adult rats in terms of the effects of RF exposure on the BBB.
The BBB is an important element in cerebral microcirculatory functions and is known to be a significant target for evaluating the adverse effects of RF exposure. Since 1994, Salford's group suggested that BBB permeability is modified by RF exposure, even at low intensities, below the restriction values established by the ICNIRP (5). Histological examination found leakage of endogenous albumin in rat brains after whole-body exposure at intensities in the milliwatt per kilogram range (22, 23). However, other research groups failed to confirm this albumin leakage under various exposure conditions, including those used in the original experiment (21, 24-26). Furthermore, this finding was also verified in developing rats, for which the debate on the impact of RF exposure on the BBB had not yet been settled (4). Kuribayashi et al. found no albumin leakage in immature, 4-week-old rat brains, even after a 2-week-exposure at an brain average SAR of 6 W/kg (13). Our findings support the results of Kuribayashi's group. In support of their findings in postmortem rats, direct observation in vivo during RF exposure revealed no change in BBB permeability. Of course, another issue is still pending: Can these findings, obtained in developing rats, be extrapolated to developing human children? However, the fact that RF exposure did not modify BBB permeability in experiments using animals in the developmental stage may be helpful for discussing physiological changes in the human brain following local exposure to RF.

Immunohistological evaluation of serum albumin leakage. Rat brains were fixed with 4% paraformaldehyde after radiofrequency electromagnetic field (RF) exposure and sections were prepared for histological evaluation. Representative images show the cortex of the target area, the hippocampus below the target area, and the brain base. No sites immunostained with antibodies to albumin were observed in either 4-week-old juvenile rats (n=3 per group) or 8-week-old young adult rats (n=3 per group). Bar: 300 μm.
In conclusion, our research explored dynamic changes in BBB permeability in juvenile and young adult rat brains during local cortex exposure to RF under non-thermal conditions. However, like many other studies using adult rats, no dynamic changes in BBB permeability related to RF were observed during 50-min exposure at 2.0 W/kg TASAR. Therefore, RF exposure below the local exposure limit (2 W/kg) is unlikely to induce dynamic or delayed effects on BBB, even at developmental stages. However, further investigations are required, as the effects of RF exposure on other cerebral microcirculatory parameters, such as hemodynamics, in the developing stage are still unclear.
Acknowledgements
This study was supported by the Ministry of Internal Affairs and Communications, Japan. The Authors thank Mrs.' Miyuki Takahashi, Kasumi Yamanaka, and Minako Segawa for their helpful assistance.
Footnotes
-
↵* These Authors contributed equally to this study.
-
Competing Interests
The Authors have declared that no competing interests exist.
- Received March 16, 2015.
- Revision received March 24, 2015.
- Accepted March 26, 2015.
- Copyright © 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}