


Abstract
Background/Aim: Nicotine (NIC) exposure – via smoking and the increasingly popular E-cigarettes- increases oxidative stress and hence, renal risk in smokers. Resveratrol (RES) may help ameliorate this risk by mounting anti-oxidant responses in the kidney. Materials and Methods: Renal proximal tubule cells (NRK52E) were treated with vehicle or 20 μM RES prior to treatment with 200 μM NIC and generation of reactive oxygen species (ROS) as well as cell viability was determined. RES-induced antioxidant responses were determined in reporter luciferase assays. Gene silencing was used to determine mechanism of RES action. Results: RES protected NRK52E cells from NIC-induced oxidative injury. RES activated the promoter of the anti-oxidant manganese superoxide dismutase (MnSOD) gene via activation of the forkhead box O (FoxO3a) transcription factor. Silencing of MnSOD abolished the protective effects of RES on NIC-associated oxidative injury. Conclusion: RES may provide protection to the kidney from the adverse effects of NIC in smokers.
Smoking –through its nicotine (NIC) content (1)– is an important risk factor in progression of kidney diseases (2). Increasing popularity of E-cigarettes (3, 4) could further increase this risk as they are believed to be safe alternatives to tobacco smoking (3). While the best remedy would be to stop smoking, many smokers refuse to quit (5). Hence, alternative means may be needed to ameliorate adverse effects of smoking/NIC-exposure on the kidney. Since smoking/NIC exposure increases oxidative stress in the kidney (6, 7) and in cultured renal proximal tubule cells (7, 8), antioxidants may fulfill this protective role. Some naturally-occurring compounds, such as resveratrol (RES), have shown antioxidant capacity (9) and protective effects of RES against smoking-associated renal tubular damage have been shown (10). Regardless, the mechanism by which RES protects the kidney is virtually unknown.
Antioxidant properties of RES are associated with its ability to induce expression of antioxidant genes including manganese superoxide dismutase (MnSOD) (11, 12). One possible mechanism by which RES upregulates MnSOD is activation of the transcription factor cAMP responsive element binding protein (CREB) (13) or Forkhead box O (FoxO3a) (11). However, this effect of RES on the kidney under NIC exposure is unknown.
Accordingly, the aim of this study was to determine whether (i) RES protects renal proximal tubule cells from NIC-mediated oxidative injury and (ii) the protective effects of RES due to activation of the CREB/FoxO/MnSOD pathway.
Materials and Methods
Cell culture and treatment. The rat proximal tubule cell line (NRK52E) was maintained in 5% CO2 at 37°C in DMEM with 10% FBS as described elsewhere (14). Some cultures were pre-treated with 20 μM resveratrol (RES, Selleckchem, Houston, TX, USA) or vehicle (DMSO) prior to treatment with 200 μM nicotine (NIC; Sigma-Aldrich, St. Louis, MOS, USA)
Determination of reactive oxygen species (ROS) production. Intracellular generation of ROS was determined by fluorescent oxidant-sensitive 2’,7’-dichlorofluorescein-diacetate dye (DCFDA; Life Technologies, Grand Island, NY, USA). Briefly, cells grown in T25 flasks and treated with 20 μM RES or vehicle overnight were collected, counted and loaded with 100 μM DCFDA for 30 min. After washing away the excess dye equal number (0.5×106) of cells were distributed in a 96-well plate; 200 μM NIC was added and ROS production was immediately determined as described elsewhere (15). ROS production was calculated as the increase in fluorescence/30 min/0.5×106 cells and expressed as a percentage of that of corresponding untreated cells.
Viability assay. For assessing cellular viability, the Cell Titer Blue fluorescent kit was used (Promega, Madison, WI, USA). Briefly, cells grown in 96-well plates were transfected/treated as needed and reagent added. After incubation for 2 h at 37°C fluorescence was determined. Results were expressed as percentage of control values.
Reporter luciferase assay. NRK52E cells grown in 24-well plates were transfected with either a MnSOD-promoter-reporter-luciferase (16), a luciferase plasmid that harbors 6 canonical FOXO binding sites (6xDBE) to determine FOXO-dependent transcription (17) or cAMP responsive element (CRE) luciferase (Stratagene, La Jolla, CA, USA) together with Renilla luciferase (Promega, Madison, WI, USA) by using Lipofectamine 3000 reagent (Life Technologies, Grand Island, NY, USA). 24 h after treatment firefly and renilla luciferase activities were determined by using the Dual Luciferase assay kit (Promega, Madison, WI, USA). Luciferase activities were calculated as the ratio of the firefly and renilla activities and expressed as a percentage of the control (untreated) values.
Knockdown of FoxO3a and MnSOD. Knockdown of FoxO3a was performed via transfection of a FoxO3a siRNA (Cell Signaling Technologies, Danvers, MA, USA) using the Lipofectamine 3000 transfection reagent (Thermofisher Scientific, Waltham, MA, USA) according to the manufacturer's recommendation. To knockdown MnSOD, NRK52E cells were transfected with a short-hairpin-MnSOD (shMnSOD) plasmid as described elsewhere (14). Knockdown of MnSOD was verified by Western blotting (14).
Statistical analysis. Continuous variables are expressed as means and standard deviations (S.D.). One-way ANOVA with Holm-Sidak post-hoc test was used to evaluate differences between groups. Differences between means were considered significant if p<0.05. All analyses were performed using the GraphPad InStat3 (La Jolla, CA, USA) software package.
Results
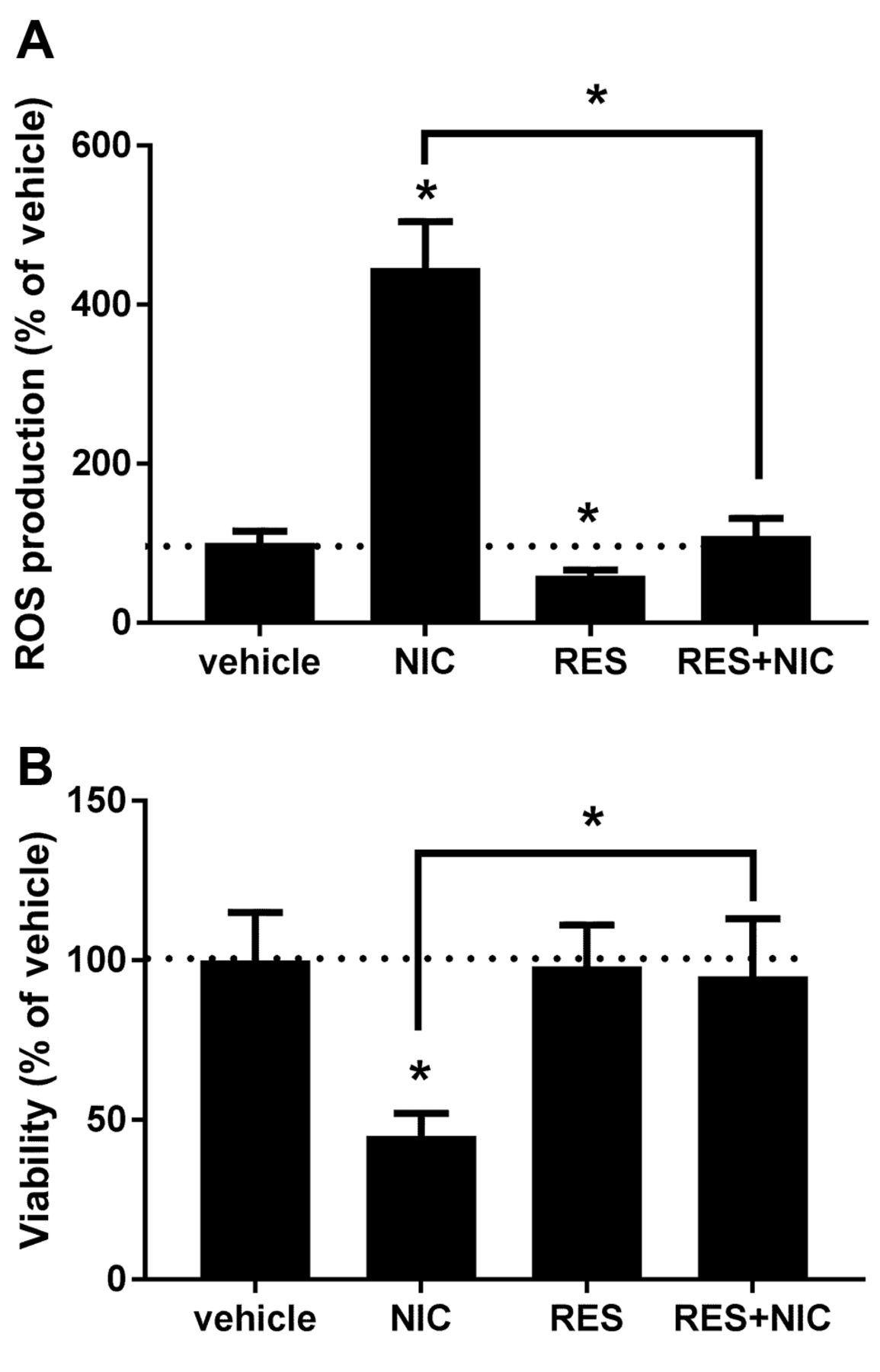
Resveratrol protects renal proximal tubule cells from nicotine-mediated oxidative stress and injury. NRK52E cells were treated with vehicle or 20 μM RES overnight and 200 μM NIC-mediated ROS production was determined as described in Materials and Methods. Figure 1A shows that pre-treatment with RES completely abolished NIC-induced ROS release. It is important to note that RES alone attenuated endogenous ROS production. Since NIC-induced ROS leads to significant cytotoxicity in renal proximal tubule cells (7), we determined whether RES protects cells from NIC-associated cytotoxicity. Accordingly, NRK52E cells were treated with vehicle or 20 μM RES for 24 h followed by treatment with 200 μM NIC. Viability of cells was determined 24 h later. Results shown in Figure 1B demonstrate that RES, indeed, protects from cytotoxicity imposed by NIC treatment.
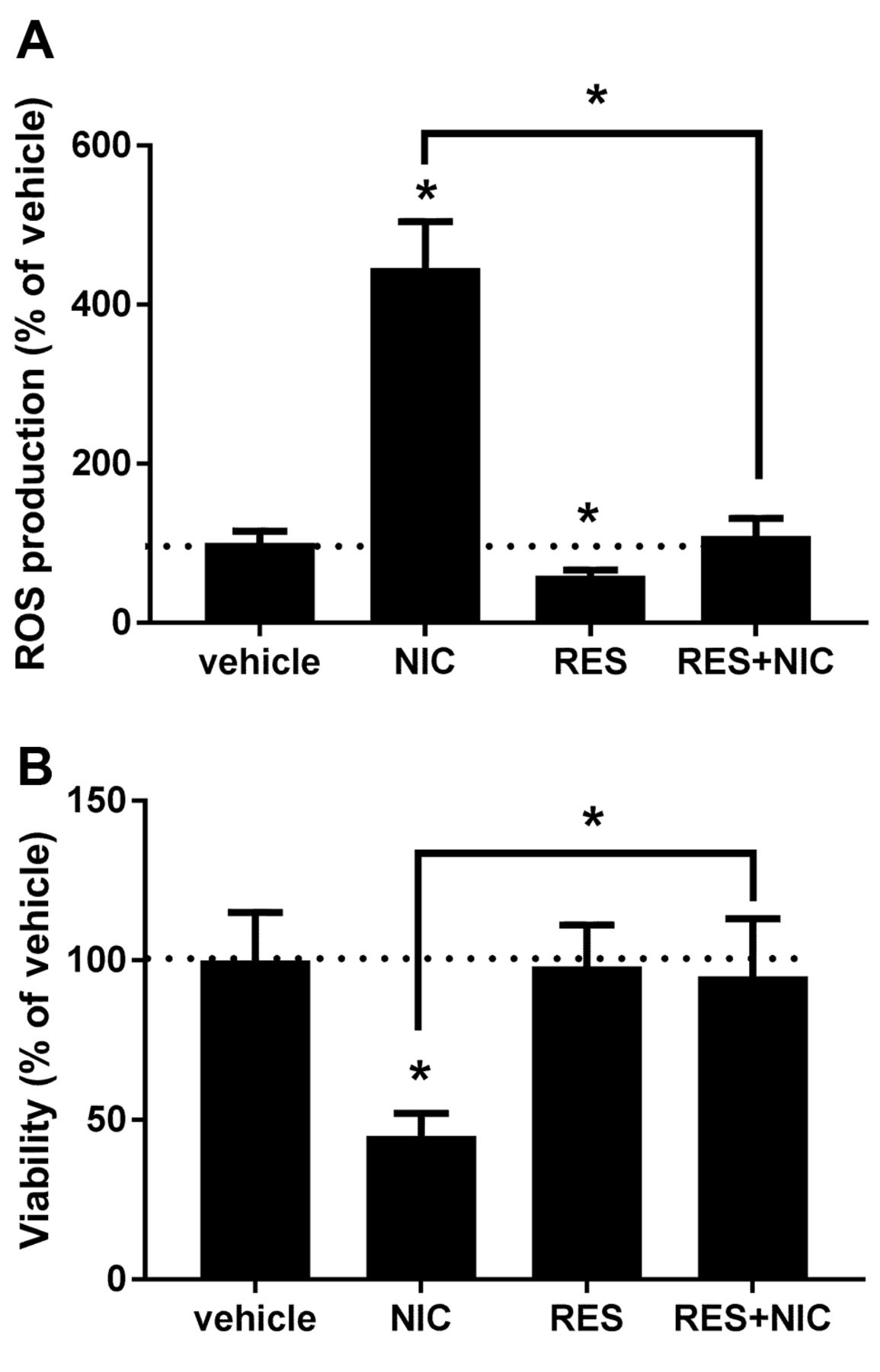
Resveratrol protects renal proximal tubule cells from nicotine-mediated oxidative injury. NRK52E cells were pretreated with vehicle or 20 μM RES for 24 h and 200 μM NIC-induced ROS production (A) as well as viability (B) was determined. n=3; *p<0.05 compared to vehicle or as indicated. The dotted line marks control (vehicle-treated) value.
Resveratrol activates the promoter of the antioxidant MnSOD gene via FoxO3a. To determine whether RES activates the antioxidant MnSOD, NRK52E cells were transfected with a reporter luciferase that harbors the promoter of the MnSOD gene (16) together with a renilla luciferase, as described in Materials and Methods. Transfected cells were then treated with 20 μM RES and 24 h later luciferase activities were determined. Figure 2A shows that RES significantly activated the MnSOD promoter. To verify that RES induces the MnSOD promoter via CREB (13) or FoxO (18) transcription factors, NRK52E cells were transfected with either a CRE or a reporter luciferase plasmid that harbors 6 canonical FOXO binding sites (6xDBE) together with Renilla luciferase. Figure 2A demonstrates that 24 h treatment with 20 μM RES significantly increased the activity of 6xDBE reporter, but not the CRE reporter. To demonstrate that FoxO plays a role in RES-mediated induction of the MnSOD promoter, NRK52E cells were co-transfected with a FoxO3a siRNA plus a MnSOD promoter luciferase and treated with 20 μM RES for 24 h. Figure 2A demonstrates that silencing FoxO3a (siFoxO3a) significantly attenuated RES-mediated induction of the MnSOD promoter. The extent of siRNA-mediated FoxO3a knockdown (~70%) is shown in Figure 2B.

Resveratrol activates the promoter of the MnSOD gene via FoxO3a. (A) NRK52E cells were transfected with either a MnSOD promoter, CRE or 6xDBE luciferase reporter together with a renilla luciferase. Some cells were co-transfected with a FoxO3a siRNA plus MnSOD/renilla plasmids. Transfected cells were treated with 20 μM RES for 24 h. n=3; *p<0.05 compared to vehicle or as indicated. The dotted line marks control (vehicle-treated) value. (B) The extent of knockdown was verified by Western blotting in NRK52E cells that were transfected with a FoxO3A siRNA (siFoxO3a). Data are representatives of 3 individual experiments. scrFoxO3a: scrambled FoxO3a.
Silencing of MnSOD attenuates the protective effects of resveratrol on nicotine-associated cytotoxicity. NRK52E cells were transfected with shMnSOD to knockdown MnSOD expression as described in Materials and Methods: extent of silencing was determined elsewhere (14). Control cells were transfected with an empty plasmid. Transfected cells were pretreated with 20 μM RES overnight and 200 μM NIC-mediated ROS production and cytotoxicity were determined. Figure 3 demonstrates that silencing of MnSOD reversed protective effects of RES on NIC-mediated ROS release. Not surprisingly, silencing of MnSOD also decreased viability of cells treated with RES+NIC.

Resveratrol protects renal proximal tubule cells from nicotine-induced cytotoxicity via MnSOD. NRK52E cells were co-transfected with shMnSOD together with MnSOD promoter reporter and treated with 20 μM RES. After 24 h 200 μM NIC-induced ROS production and cell viability were determined as described in Materials and Methods. n=3; *p<0.05; upper and lower dotted lines mark NIC values for ROS and viability (from Figure 1), respectively.
Discussion
It is well accepted that smoking/NIC exposure increases oxidative stress in the kidney that enhances the risk for development and progression of renal chronic diseases (1, 6-8). NIC-associated oxidative stress is related to release of excessive amount of ROS from the mitochondria (7, 8, 19), which may not be counterbalanced by the resident antioxidant system such as the mitochondrial MnSOD (20). Therefore, modalities that reinforce the endogenous antioxidant system would be beneficial in ameliorating smoking/NIC exposure-associated oxidative stress.
Resveratrol (RES) is a naturally occurring antioxidant, found in high concentration in grapes and wine (21). Importantly, RES activates mitochondrial antioxidant enzymes, such as MnSOD, that decrease mitochondrial ROS production (22). Thus, RES is a good candidate to prevent smoking-associated oxidative stress. Indeed, several studies have shown beneficial effects of RES on smoking in various organs (22-24), while the kidney is barely studied (10). Our study demonstrated that RES protected rat renal proximal tubule cells from NIC-mediated oxidative stress (Figure 1B) by reducing ROS production (Figure 1A). We also found that this protection is attributed to the activation of MnSOD (Figure 3).
Transcriptional regulation of MnSOD is associated with induction of several transcription factors that bind to the promoter of the MnSOD gene. One candidate is the CREB transcription factor (13), which is actually activated by RES in the heart (25) and brain (26). In our system, we did not find significant induction of a CRE luciferase reporter after treatment with RES (Figure 2). Besides CREB, FOXO transcription factors are also involved in activation of the MnSOD promoter (27). Renal proximal tubule cells express FoxO3a (28) and earlier we showed that FoxO3a is involved in MnSOD induction in renal proximal tubule cells (14). Studies have also shown that RES activates FoxO3a in kidney cells (11). Our study confirmed this hypothesis: RES treatment significantly induced FOXO-dependent transcription (Figure 2). We also demonstrated that FoxO3a mediates RES-dependent induction of the MnSOD promoter (Figure 2).
In conclusion, RES attenuates NIC exposure-associated induction of ROS release and consequent renal toxicity via FoxO3a-dependent induction of the antioxidant MnSOD gene. Our results showed that treatment with RES may be used to ameliorate smoking/NIC exposure-inflicted renal risk.
Acknowledgements
This work was supported by a grant from the Department of Pediatrics at the University of Mississippi Medical Center and the Bower Foundation.
Footnotes
This article is freely accessible online.
- Received March 23, 2017.
- Revision received April 6, 2017.
- Accepted April 7, 2017.
- Copyright© 2017, International Institute of Anticancer Research (Dr. George J. Delinasios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.