


Abstract
Background: Mitochondrial DNA (mtDNA) encodes for the respiratory chain proteins. Genetic alterations in mtDNA have been described during aging and linked to impaired hematopoiesis. Materials and Methods: We investigated two novel conplastic mouse strains harboring a mitochondrial nt7778 G/T polymorphism leading to an amino acid exchange in respiratory chain complex V. Effects on reactive oxygen species (ROS) and adenosine triphosphate (ATP) levels, as well as bone marrow composition and peripheral blood counts, were investigated during aging (up to 24 month). Results: The polymorphism correlated with significantly decreased ROS levels in aged mice. Effects on hematopoiesis were marginal and not statistically significant: numbers of erythroid cells in bone marrow, as well as mean corpuscular hemoglobin, tended to decrease over time. Conclusion: The investigated polymorphism is associated with decreased ROS levels in aged hematopoietic cells but does not significantly influence hematopoiesis itself.
Aging of the hematopoietic system is accompanied by several changes in different blood compartments. For example, an immune shift in aged individuals has been described, leading to a higher percentage of myeloid cells with a concomitant enhanced susceptibility to infections and development of anemia (1, 2). Clinically, elderly individuals are more affected by myelodysplastic syndromes (MDS) and leukemia (3).
At the subcellular level, several modifications have been described during aging in human hematopoietic stem cells (HSCs). Elevated levels of phoshorylated histone H2AX (gammaH2AX), correlating with DNA damage, were found in HSCs of older individuals in comparison to HSCs of the umbilical cord blood (4). A transgenic mouse model of MDS showed increased levels of reactive oxygen species (ROS), correlating with increased levels of DNA damage and a more intensive progression from MDS to acute myelogenous leukemia (5). Further mouse models investigated the influence of manipulated mitochondrial DNA (mtDNA) on hematopoiesis. They revealed a correlation between impaired mtDNA function, elevated ROS levels and deregulated HSC function (6-8).
The mtDNA encodes for 13 proteins that are all part of the respiratory chain complexes (9). The respiratory chain generates a proton gradient (complex I, III and IV), allowing the respiratory chain complex V (FOF1-ATPase) to synthesize adenosine triphosphate (ATP). Besides ATP synthesis, the complexes of the respiratory chain are also the main source for ROS in the cell. ROS molecules as superoxide or hydrogen peroxide (H2O2) are highly reactive and, at high concentrations, able to cause serious damage to cell components, such as double-strand breaks (10) and membrane disintegration (11, 12). Moreover, it was shown that elevated ROS levels lead to reduced maintenance and repopulation capacity of murine hematopoietic stem cells (HSC) (13).
Nevertheless, H2O2 carries out important positive functions in the cell as increased scavenging of ROS by overexpression of antioxidants leads to decreased proliferation by inactivation of Akt (14). Several signaling pathways are described to be activated by ROS generation, including NF-κB, PI3K, MAPK and p53 (15). Furthermore, H2O2 stimulation induces proliferation of mammalian cells (14, 16, 17). It was also shown that signals stimulating hematopoietic growth factors are transduced by ROS formation (18). However, the link between ROS, mtDNA mutations, hematopoietic aging and hematological diseases are not yet completely understood.
Here, two conplastic mouse strains differing in one single polymorphism within the mtDNA ATP8 gene were comparatively analyzed. In the respiratory complex V, the ATP8 protein is part of the proton channeling FO subunit. The C57BL/6-mtFVB/NJ (mtFVB) strain harbors the nt7778G/T polymorphism, while the control C57BL/6-mtAKR/J (mtAKR) strain is unaltered at this position. The replacement of aspartic acid, which lacks an aromatic group with tyrosine at the fifth position of the highly-conserved N-terminal region, is important for proper complex V assembly (19). Due to the proton channeling function of subunit FO, we hypothesize that the polymorphism will alter the transport of protons by complex V and, therefore, the generation of the proton gradient. It would, thus, impact the whole respiratory chain, influencing ROS and ATP production. Recently, the impact of the nt7778 G/T polymorphism on cell function has been studied in different tissues (19-22), as well as its effects on anxiety and stress reactivity (23, 24).
We analyzed the effects of this polymorphism on the hematopoietic system, particularly in hematopoietic stem cell function during aging. Groups of both strains at three different time points ranging from young (3 months), mid-aged (12 months) to advanced-aged (24 months) were included in the study. In addition to analyzing the blood count at these time points, we also investigated the ROS and ATP levels of bone marrow cells (BMC), as well as the proportion of distinct subpopulations. Furthermore, an in vivo study was performed to investigate the restoration of hematopoiesis after cytostatic stress. This revealed a correlation between the nt7778 G/T polymorphism and the aging process in murine hematopoiesis.
Materials and Methods
Animals. Conplastic mouse strains were implemented as described before (24). In brief, due to the maternal inheritance of mtDNA, female animals of the mitochondrial donor strain AKR/J and FVB/NJ, respectively, were crossed with males with the corresponding genomic background (C57BL/6NTac). Female offspring was then bred with males of the background strain. After ten generations the conplastic mouse strain harbored a stable mtDNA of the donor strain, as well as a stable nucleic DNA of the background strain. Animals were bred and housed under specific pathogen-free conditions in the central animal facility of the University of Rostock, Germany. Animals were kept under a 12-hour light/dark cycle and had access to water and standard laboratory chow ad libitum. All animals received human care according to the German legislation on protection of animals and the Guide for the Care and Use of Laboratory Animals (NIH publication 86-23, revised 1985); all efforts were made to minimize suffering. For in vitro analysis of BMC, male and female animals at 3, 12 and 24 months of age were investigated. For in vivo study, animals at 3 and 12 months were examined. Animal experiments were approved by the state's Animal Care Committee of Mecklenburg-Western Pomerania (LALLF M V/TSD/7221.3-1.1-100/12).
Isolation of BMC. Animals were sacrificed under narcosis (ketamine 65 mg/kg of body weight and xylazine 13 mg/kg of body weight) by cervical dislocation. Tissue was removed from femora and tibiae, which were cut at both sides and flushed several times with PBS. Cells were collected in falcon tubes and kept on ice until further processed.
Blood count. Blood samples were obtained from the retro-orbital plexus using an ethylenediaminetetraacetic acid (EDTA)-coated capillary and collected in EDTA-coated tubes (Sarstedt, Nümbrecht, Germany). Samples were diluted 4-fold with saline for analysis on an Advia 2120 hematology analyzer (Siemens Healthcare, Erlangen, Germany). For in vivo study, peripheral blood (50 μl) was collected every 2 to 4 days by tail bleeding up to day 15 and 21, respectively.
Measurements of ROS and ATP levels. For measurement of complete ROS, BMC were incubated with 50 μM 2’,7’-dichlorofluorescein diacetate ((DCFH-DA); Sigma, Taufkirchen, Germany) for 30 min. Fluorescence of 5×104 cells was measured using a Glomax® microplate reader (Promega, Mannheim, Germany) at a wavelength of 485 nm.
To investigate mitochondrial superoxide, cells were stained with MitoSOX™ (Invitrogen, Darmstadt, Germany) and Annexin-V-APC (Beckton Dickinson, Heidelberg, Germany) to exclude apoptotic cells from measurement. Cells were analyzed with a FACSCalibur (Beckton Dickinson, Heidelberg, Germany) and the proportion of MitoSOX™-positive and Annexin-V-APC-negative cells was counted.
Intracellular ATP levels of BMC were analyzed using the ATP Bioluminescence Assay Kit HS II (Roche, Mannheim, Germany). In brief, luminescence of cell lysates of 5×104 cells mixed with luciferase reagent was measured with a Glomax® reader. By using a standard serial dilution of ATP, the concentration in cell lysates could be determined.
Immunophenotyping. To evaluate the amount of the different subpopulations in the bone marrow, cells were stained with fluorescent antibodies (all obtained from Beckton Dickinson, Heidelberg, Germany) against different surface antigens: lymphocytes (IgG2 anti-CD3ε phycoerythrin (PE) (Cat.-No. 553240), IgG2a anti-CD45R fluorescein isothiocyanate (FITC) (Cat. No. 553087)); erythroid cells (IgG2b anti-Ter-119 allophycocyanin (APC) (Cat.-No. 557909), IgG1 anti-CD71 PE (Cat.-No. 553267)); stem cells (IgG2b anti-c-kit PE (Cat.-No. 553355), IgG2a anti-Sca-1 PE-cyanine 7 (Cy7) (Cat.-No. 558162), lineage cocktail APC (Cat.-No. 558074), IgG2a anti-CD34 FITC (Cat.-No. 560238)) and corresponding isotype controls (hamster-IgG2 PE (Cat.-No. 550085), iso-lineage cocktail APC (Cat.-No. 558074), rat-IgG2a FITC (Cat.-No. 553929), rat-IgG2a PE-Cy7 (Cat.-No. 557855), rat-IgG2b PE (Cat.-No. 556925), rat-IgG1 PE (Cat.-No. 554685), rat-IgG2a FITC (Cat.-No. 553929)). Stained cells were analyzed with FACSCalibur (Beckton Dickinson). Total nucleated cells (TNC) were gated and the amount of cells positive for the distinct antigens was evaluated.

Polymorphism nt7778 G/T correlated with varying reactive oxygen species (ROS) and adenosine triphosphate (ATP) levels. (A) Total intracellular ROS in bone marrow cells was determined by staining with 2’,7’-dichlorofluorescein diacetate (DCFH-DA) and measuring the relative fluorescence units (RFU). ROS levels are displayed for the three analyzed aging stages of both conplastic strains. (B) Bone marrow cells containing mitochondrial superoxide were stained with MitoSOX™ and analyzed by flow cytometry. Additionally, the cells were stained with Annexin-V-APC to exclude apoptotic cells with unspecific signals. (C) ATP levels were evaluated by adding luciferase to cell lysates and measuring luminescence. *Significant difference versus mtAKR strain at same aging stage. Brackets indicate significant changes within one strain during aging. Six or more mice were investigated for each strain and aging stage.
In vivo studies. The reconstitution capacity of the hematopoietic system was investigated by cytostatic stress induction. Animals were kept in a specific pathogen-free environment. After initial blood collection, animals were injected intraperitoneally with 150 mg/kg of body weight 5-fluorouracil (5-FU). The corresponding control group of each strain was treated with an equal volume of saline. For blood counts, whole blood (50 μl) was taken every 2 to 4 days by tail bleeding up to days 15 and 21, respectively. After initially 21 days, bone marrow was isolated and investigated as described above. Later, bone marrow was analyzed at day 15 (12-month-old animals: n=5, 3-month-old animals: mtAKR: n=3; mtFVB: n=4). Furthermore, spleen weight was measured. Data of the 5-FU-treated groups were analyzed in relation to the corresponding control group by calculating ratios. Study groups consisted of 10 mice.
Statistical analysis. Results within each experiment were described using mean and standard deviation. Significance between strains was calculated using Mann-Whitney U-test (SPSS Software, Version 22; IBM, Ehningen, Germany). A p-value <0.05 was considered to be significant.
Results
Polymorphism nt7778 G/T correlated with changes in ROS and ATP levels in aged mice. The ROS levels were measured by two different methods. Total cellular ROS was determined by DCFH-DA staining (Figure 1A) and the cells containing mitochondrial superoxide were analyzed by using MitoSOX™ (Figure 1B). While control mtAKR animals showed a slight increase of intracellular ROS from 3 to 24 months (7.1±1.7×103 relative fluorescence units (RFU) to 9.0±4.0×103 RFU; p=0.628), the mtFVB strain displayed a significant decrease (5.5±2.6×103 RFU to 2.4±0.7×103 RFU; p=0.037) over time. In comparison to the mtAKR strain, intracellular ROS levels at 24 months were significantly lower in mtFVB strain (p<0.001).
Concerning mitochondrial superoxide, the mtAKR strain demonstrated an non-significant increase from 0.9±0.4% of MitoSOX™-positive cells in bone marrow at 3 months to 1.5±1.1% at 24 months (p=0.414). The mtFVB strain showed almost stable levels with 0.4±0.3% in young animals and 0.5±0.4% in advanced-aged mice (p=0.323). The superoxide levels at 3 and 24 months were significantly lower in mtFVB strain compared to mtAKR (p=0.015 and p=0.030, respectively). ATP levels changed in significantly over time (Figure 1C). ATP levels were 1.0±0.7 μM in young animals and 0.4±0.1 μM in mtAKR mice of advanced age (Figure 1C; p=0.867). In comparison, the mtFVB strain displayed levels from 0.2±0.1 μM to 0.5±0.2 μM (p=0.216).
In summary, the mtFVB strain displayed significantly lower levels of ROS and mitochondrial superoxide in elderly animals, while ATP levels were unaffected.
Polymorphism nt7778 G/T correlated with differing composition of the bone marrow during aging. The proportions of cellular subpopulations in the bone marrow changed during aging in both strains (Figure 2). The LSK cells were defined as lineage, Sca-1+ and c-kit+. The percentage of LSK cells within the lineage-cells increased not significantly from 3 to 24 months (mtAKR: 2.1±0.7 to 2.8±1.1%; p=0.345; mtFVB: 2.1±1.1 to 3.5±1.6%; p=0.106). However, the proportion of CD34dim/− cells in LSK pool increased significantly from 3 to 24 months in the mtFVB strain (72.0±4.8 to 88.2±7.5%; p<0.001) and showed significantly higher levels compared to mtAKR mice at 24 months (71.8±19.2%; p=0.010). Both, the mtAKR, as well as the mtFVB strain, demonstrated a marginal increase of erythroid cells during aging (25.2±3.5 to 26.0±7.8% of TNC; p=0.189 (mtAKR); (25.9±7.3 to 18.9±7.2%; p=0.149 (mtFVB)).
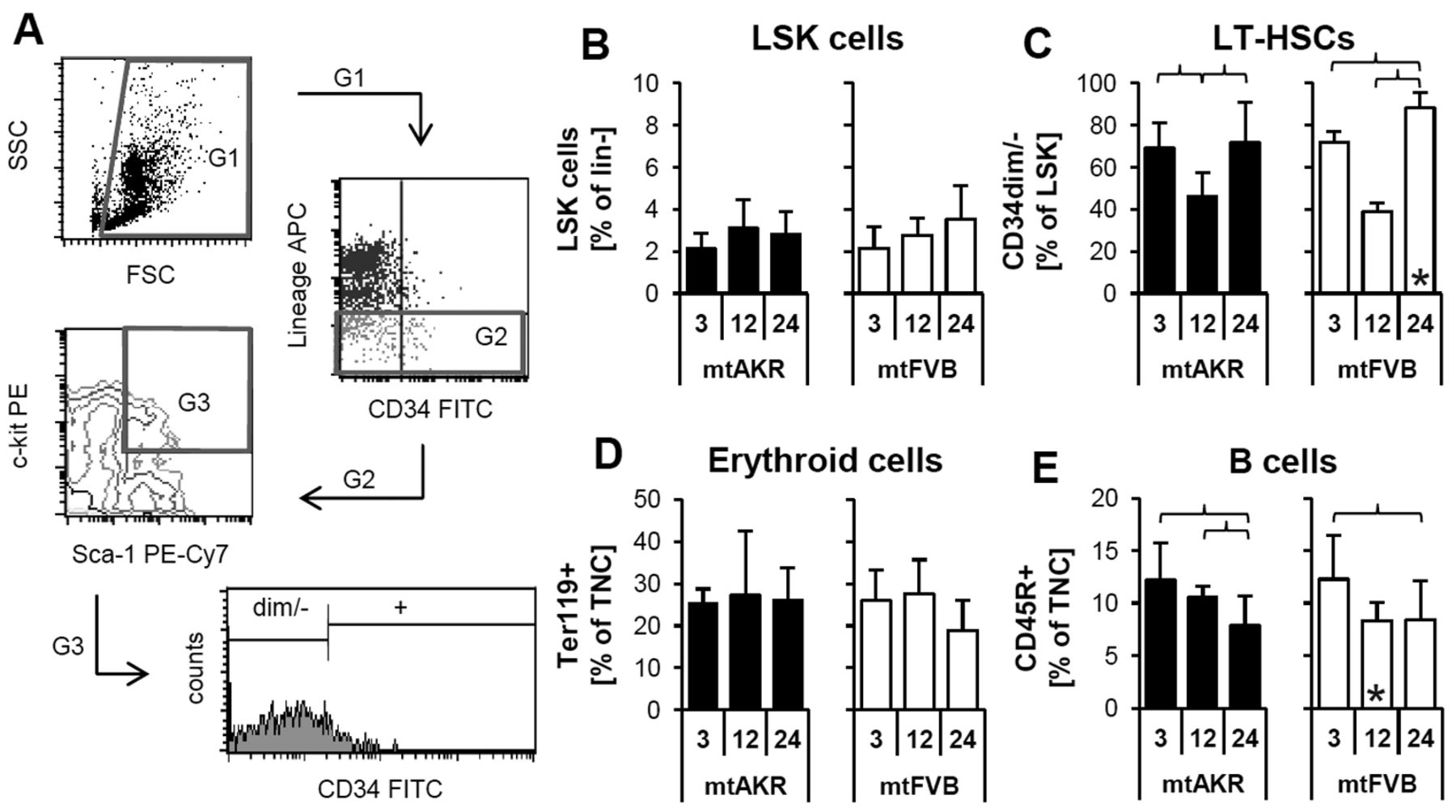
Polymorphism nt7778 G/T impacted the composition of bone marrow subpopulations. (A) Determination of LSK and LT-HSCs is shown as an example. Of the total nucleated cells ((TNC); G1), those negative for lineage commitment were determined (G2). Of these, cells positive for Sca-1 and c-kit were gated (G3; LSK cells shown in B). Subsequently, the proportion of cells with a diminished or no expression for CD34 was identified (LT-HSCs shown in C). Further, the amount of Ter119+ (erythroide cells – D) and CD45R+ (B cells – E) cells, respectively, was determined. *Significant difference versus mtAKR strain at same aging stage. Brackets indicate significant changes within one strain during aging. Six or more mice were investigated for each strain and aging stage.
Both strains exhibited a similar composition of lymphoid subpopulations. A slight increase in T cell numbers (data not shown) was detected, while B cells decreased in both strains during aging. Noteworthy, the mtFVB strain revealed an earlier decrease of B cells in mid-aged mice with a significant lower proportion of B cells (8.4±1.7% of TNCs) compared to mtAKR strain (10.6±1.1%; p=0.013).
To sum-up, the bone marrow composition of 24-month-old animals from the mtFVB strain differs significantly from mtAKR strain in the proportion of CD34dim/− cells. They also tended to have fewer erythroid cells (p=0.129). In addition, B cell populations diminished at an earlier time point in mtFVB strain compared to mtAKR strain during aging.
Polymorphism nt7778 G/T correlated with reduced hemoglobin content of erythrocytes. Blood count analysis is an important parameter to evaluate hematopoietic stem cell function. In young and mid-aged mice, both strains displayed very similar parameter patterns (Figure 3). Mice of advanced age, however, showed distinct differences. Twenty-four-month-old mice of the mtAKR strain tended to have more leukocytes (white blood cells (WBC)) compared to the mtFVB strain (6.2±3.3×103/μl vs. 4.2±2.4×103/μl; p=0.098). Although the mtFVB strain displayed similar numbers of erythrocytes compared to the mtAKR samples, the hemoglobin levels (HGB) were slightly lower at 24 months compared to the control strain (10.8±1.7 μg/μl vs. 12.1±1.4 μg/μl; p=0.060). The mean corpuscular hemoglobin (MCH) exhibited significantly lower levels in mtFVB mice than in the mtAKR strain at 24 months (12.2±1.4 vs. 14.1±1.7 pg/cell; p=0.003). Levels of both HGB and MCH in mtFVB animals were below the murine standard (HGB: 11.8-14.9 μg/μl; MCH: 13.8-18.4 pg/cell). Furthermore, in 24-month-old animals, the proportion of neutrophils (the most abundant type of granulocytes of total WBCs) was marginally higher in mtFVB strain with 8.6±3.6% than in the mtAKR strain with 6.0±2.0% (p=0.059). Lymphocyte levels were not significantly different between the strains.
In summary, older mtFVB animals displayed a more distinct age-related decrease of HGB and MCH compared to mtAKR.
Polymorphism nt7778 G/T correlated with altered restoration of hematopoietic system after cytostatic stress in vivo. Following treatment with 5-FU, both strains developed a pancytopenia with subsequent recovery of blood parameters. The results of the 5-FU-treated animals were analyzed in relation to the data of the corresponding control group treated with saline solution by calculating ratios. Analysis of WBC, red blood count (RBC), hematocrit (HCT) and platelets (PLT) revealed similar patterns in both strains (Figure 4). Both strains showed a decrease of neutrophils after cytostatic stress with a subsequent rebound reaction (Figure 5). The mtAKR strain reached the nadir at day 6 at both aging stages with a ratio of 0.12±0.13 in young mice and 0.07±0.05 (Figure 5B) in older animals. Notably, young animals of the mtFVB strain also displayed the nadir at day 6 but with a higher ratio of 0.23±0.14 (p=0.053). Mid-aged mice of mtFVB strain possessed more neutrophils at day 6 with a ratio of 0.20±0.14; however, this difference did not reach statistical significance (p=0.063). They presented their nadir at day 9 with a ratio of 0.13±0.10. The rebound reaction of neutrophils in young animals from the mtFVB strain at day 15 was significantly higher with 2.51±1.23 vs. 1.21±0.53 in mtAKR animals (p=0.013). The latter displayed the restoration peak at day 18 with a ratio of 1.40±0.38. Mid-aged mice of both strains showed a similar rebound reaction in neutrophils following cytostatic stress with a ratio of 2.65±1.66 in mtAKR and 2.72±0.69 in mtFVB strain at day 15, respectively (p=0.780).

Polymorphism nt7778 G/T affected hemoglobin content of erythrocytes. Blood parameters were measured with the Advia® device. (A) Results at the three investigated aging stages (3, 12 and 24 months) are displayed for leukocytes (WBC), hematocrit (HCT), hemoglobin (HGB) and mean corpuscular hemoglobin (MCH). (B) The composition of the leukocyte pool was investigated by determining the absolute (abs) and relative (rel) parameters of lymphocytes and neutrophils. *Significant difference versus mtAKR strain at same aging stage. Brackets indicate significant changes within one strain during aging. Seven or more mice were investigated for each strain and aging stage.
The number of large unstained cells (LUC) was reduced after cytostatic application and demonstrated a rebound reaction also in both strains. Young mice of the mtFVB strain showed a lower nadir of LUC with 0.14±0.44 at day 9 compared to the mtAKR strain with 0.33±0.43 at day 2. Interestingly, in young animals, the peak of the rebound reaction seemed higher (2.15±1.02 in mtFVB vs. 1.40±0.93 in mtAKR, both at day 15); however, this difference was not significant (p=0.779). Mid-aged mice displayed a more distinct depletion of LUC in the mtAKR strain with a ratio of 0.00±0.00 at day 9 compared to 0.14±0.45 in mtFVB animals at day 6. Again, the restoration peak tended to be higher in mtFVB than in mtAKR with a ratio of 8.40±7.85 vs. 3.33±1.57 (p=0.095).
The investigation of spleen and bone marrow at days 15 and 21, respectively, revealed no significant differences between both strains when treated with 5-FU. At day 15, both strains displayed elevated levels of LSK cells and a splenomegaly, which was slightly more distinct in mtFVB strain (Figure 6). Other bone marrow subpopulations exhibited decreased levels compared to the corresponding control, while ROS and ATP levels did not show distinct deviations from control group (Figure 6). Analysis at day 21, after 5-FU treatment, revealed a normal distribution of subpopulations and almost normal spleen weight in both strains (data not shown).
Summarizing, the mtFVB strain displayed a similar regeneration of blood count after 5-FU treatment, except neutrophils and LUC, compared to mtAKR. Young animals of mtFVB exhibited a significantly stronger rebound regeneration of neutrophils than mtAKR. Further, 12-month-old mtFVB mice showed a trend to a stronger rebound reaction in LUC compared to mtAKR mice. Investigation of bone marrow at days 15 and 21 did not reveal considerable differences between the strains.

Depletion and restoration of further blood parameters following cytostatic stress. Animals were injected with 5-fluorouracil (5-FU) (150 mg/kg of body weight) at day 0. Every 2-4 days, blood was taken by tail bleeding. Blood parameters were measured with the Advia® device. Data of 5-FU-treated animals are displayed in relation to the results of corresponding control group treated with saline. WBC, White blood cells; HCT, hematocrit; PLT, platelets; RBC, red blood cells.
Discussion
Conplastic mouse strains represent a suitable model to evaluate the impact of a single mitochondrial polymorphism on organs, tissues and the whole organism. In this study, we investigated the influence of the nt7778 G/T polymorphism on the hematopoietic system. This polymorphism in ATP8 gene causes an amino acid exchange from aspartic acid to tyrosine in the corresponding protein of respiratory chain complex V. Here, ROS and ATP levels, as well as subpopulations in BMC, were analyzed during aging; also, blood count was investigated. In addition, an in vivo study evaluated the restoration capacity of the hematopoietic system after cytostatic stress.
The nt7778 G/T polymorphism correlated with changes in ROS levels in animals of advanced age. The mtFVB strain, harboring the amino acid exchanging polymorphism, showed a more distinct but not a significantly different increase of LSK cells from young to advanced-aged animals compared to the reference mtAKR. While aging HSCs were shown to increase with time (2, 25), it was indicated that elevated ROS levels in C57BL/6 mice led to an attenuated HSC maintenance and repopulation capacity (13). Furthermore, a more quiescent behavior of HSCs with higher maintenance capacity and less cycling was linked to low intracellular ROS levels (26). Here, mtFVB strain displayed a significantly higher proportion of CD34dim/− cells in LSK pool. The lack of CD34 was shown to be characteristic for LT-HSCs in mice (27). The observation of lower ROS levels in old mtFVB mice with concomitantly higher proportion of CD34dim/− cells in a larger LSK pool is in line with the already described correlation between low ROS and higher maintenance capacity of HSCs (26).
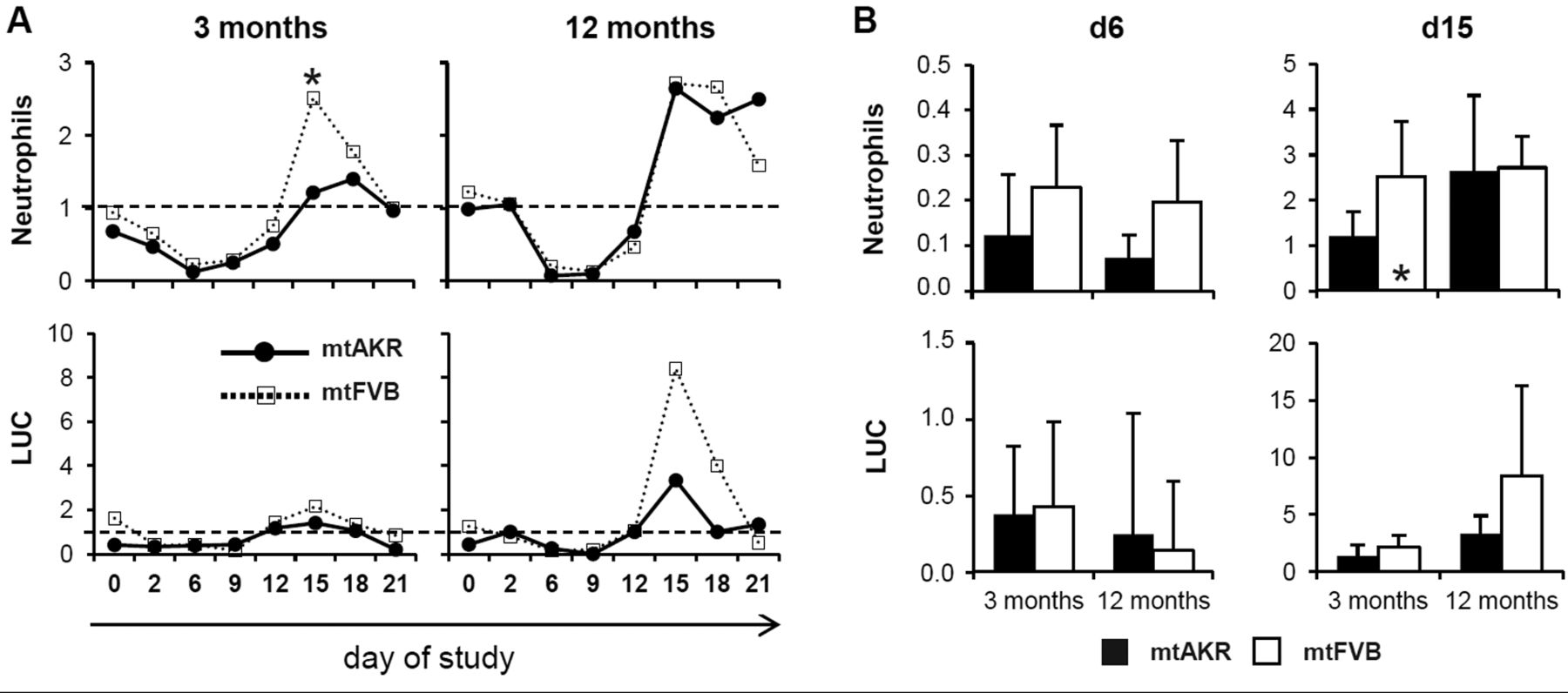
Polymorphism nt7778 G/T impacted restoration of hematopoietic system after cytostatic stress in vivo. Animals were injected intraperitoneally with 5-fluorouracil (5-FU) (150 mg/kg of body weight) at day 0. Every 2-4 days, blood was taken by tail bleeding. Blood parameters were measured with Advia®. Data of 5-FU-treated animals are displayed in relation to the results of the corresponding control group treated with saline. (A) Development of neutrophils and large unstained cells (LUC) after 5-FU treatment. (B) Neutrophils and LUCs are directly compared at days 6 and 15. *Significant difference with p<0.05.
Interestingly, the investigation of pancreatic and hepatic cells of the mtFVB strain by other groups revealed enhanced ROS levels compared to controls. Pancreatic islet cells in 3-month-old mtFVB mice showed an increased basal production of ROS compared to mtAKR strain. At the same time a pancreatic β cell dysfunction, as well as an altered mitochondrial morphology in spleen cells, could be demonstrated (21). Pancreatic acini of 12-month-old mtFVB mice, by trend, had also enhanced ROS levels in comparison to mtAKR mice (20). However, in a corresponding in vivo study, no aggravation of acute pancreatitis in mtFVB strain was observed. The induction of an acute endotoxemic liver failure revealed a survival advantage of mtFVB strain, although here oxidative stress was also enhanced compared to the background strain C57BL/6J (22).
Yu et al. also investigated the nt7778 G/T polymorphism but with different background strains that were all associated with autoimmune diseases. They reported a higher ROS production in spleen cells but no impaired mitochondrial respiration in 3- to 6-month-old animals (19). These findings differ from ours as we observed lower levels of ROS in BMC in the mtFVB strain. Therefore, the data suggest that murine tissues are differently impacted by the polymorphism concerning the formation of ROS. This could be due to the fact that the activation of distinct signaling pathways varies in different tissues, particularly under the influence of additional challenges, such as autoimmune stress. Given the participation of ROS in signaling, a varying ROS production, based upon tissue-specific pathway activation, appears plausible. Furthermore, different cell types were shown to posses varying activation of the respiratory chain with concomitantly differing ROS production. LT-HSCs, for example, were shown to cover their energy requirements by glycolysis instead of oxidative phosphorylation (28).
At the different aging stages studied here, complete intracellular ROS and mitochondrial superoxide did not change uniformly. Differences in complete ROS and superoxide can be explained by elevated degradation of superoxide to H2O2 by superoxide dismutases (SODs). Additionally, there are several enzymes in the peroxisome, as acyl-CoA oxidases, producing directly H2O2 independently from superoxide (29).
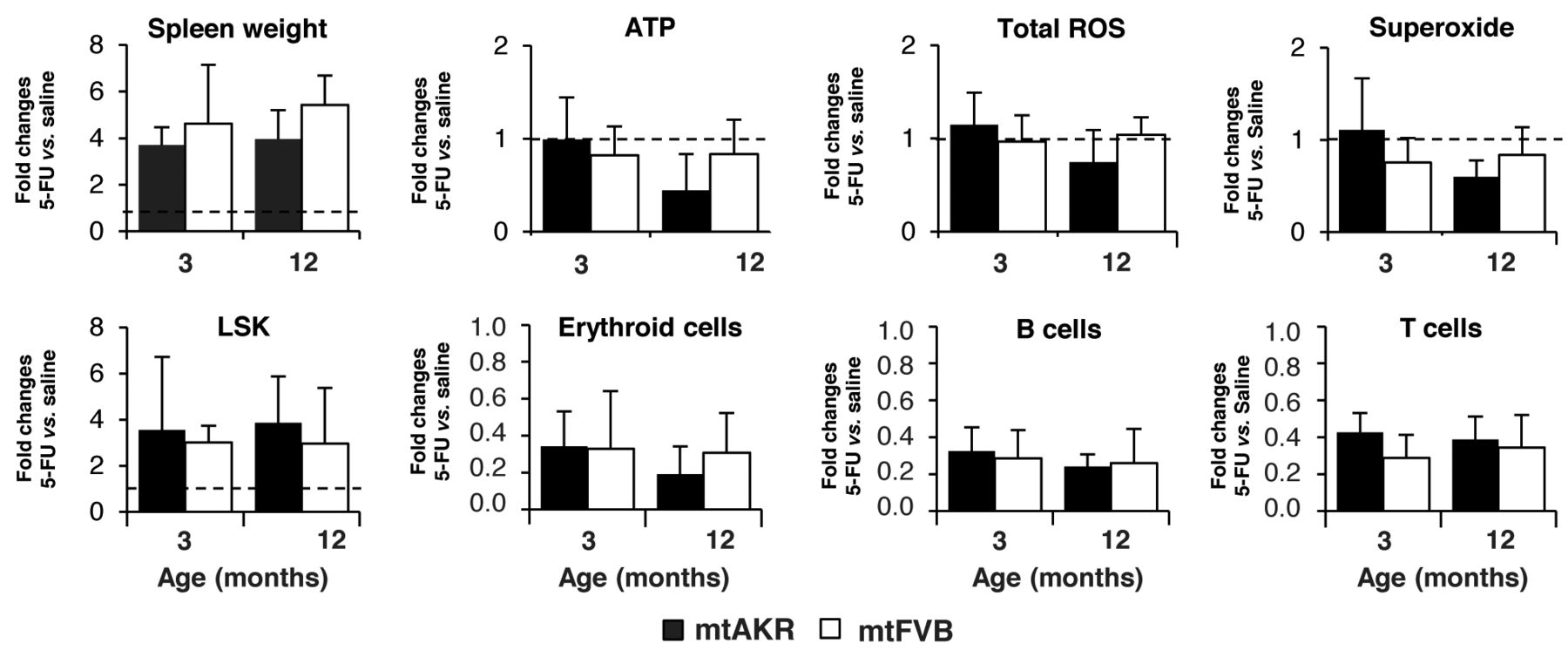
Spleen weight, reactive oxygen species (ROS) and adenosine triphosphate (ATP) levels and composition of bone marrow at day 15 post-5-fluorouracil (5-FU) treatment. Spleen weight (ratio of spleen weight to body weight) of 5-FU-treated animals was determined and analyzed in relation to the weights of corresponding control groups treated with saline. Results for ROS and ATP levels, as well as composition of bone marrow cells, are shown at day 15. The results include data of 12-month-old animals (n=5 for each strain) and 3-month-old mice (mtAKR: n=3; mtFVB: n=4).
Numazawa et al. (30) pointed out that, in C57BL/6 mice, the administration of 5-FU in hemotoxic doses led to an induction of heme oxygenase 1, a known biomarker for oxidative stress, as well as a reduction of glutathion. Moreover, when keeping the glutathion pool constant by N-acetylcysteine treatment, the 5-FU-induced hemotoxicity could be reduced, suggesting that ROS scavenging attenuated hemotoxic effects of 5-FU (30). These observations, combined with our findings of lower ROS in aged mice, lead to the assumption that if mtFVB mice would be protected from 5-FU-induced oxidative stress, the hemotoxicity should be less distinct. However, in our in vivo study, the hemotoxic effects on WBC, HCT and PLT were similar compared to the mtAKR strain. Solely, an impact on the peak recovery of neutrophils, as well as the LUC, was detectable. Basal total ROS levels showed similar values in 3- and 12-month-old mice of both strains and only a significant difference in animals of advanced age. In line with the observation of Ito et al. that enhanced ROS levels correlate with impaired HSC repopulation potential (13), the overall regeneration capacity, in our study, is similar in both strains in 3- and 12-month-old mice.
Interestingly, a higher, though marginal, increase of LUC proportion in peripheral blood in 12-month-old mtFVB mice at day 15 following 5-FU was detected, while the LSK proportion in the bone marrow was almost the same. The LUC fraction, described as large peroxidase-negative cells, also contains, apart from enlarged lymphocytes and virocytes, stem cells (31). LUC have been studied in the diagnosis and monitoring of acute leukemias and MDS (32). A recent study revealed a discriminating power of LUC in the investigation of acute promyelocytic leukemia (33). Furthermore, it was shown that an increase of LUC in peripheral blood during chemotherapy-induced neutropenia correlates with CD34+ cells and blasts in pre-nadir and post-nadir phases (34). Moreover, an enlarged LUC population was also detected in other diseases like viral infections, chronic renal failure and malaria (35). Mice treated with 5-FU, in our study, were kept in a low-germ environment and had no morbidities prior to treatment. Therefore, a possible explanation for the enlarged LUC population in the peripheral blood of mtFVB strain could be a higher stem cell mobilization. There are several studies investigating the influence of aging on stem cell mobilization. Xing et al. (36) performed a comparative assay in young and old mice investigating the influence of granulocyte-colony stimulating factor (G-CSF) on the mobilization of colony forming cells (CFC). A three-fold higher mobilization capacity of stem cells into the peripheral blood was observed in old animals, while no differences between young and aged mice were detectable in the bone marrow (36). Further, an in vitro assay showed a two-fold reduced adhesion to a cell monolayer in old compared to young hematopoietic progenitor cells (HPC) (36).
Another study reported a two-fold decrease in relative short-term homing capacities of old HSC when co-injected with young cells into irradiated recipient mice (37). Liang et al. also discovered an age-related decline in homing efficiency in murine HSC (38). In cancer patients, with standard dose chemotherapy, a highly significant positive correlation between the amount of LUC and CD34+/CD45+ cells in the peripheral blood was observed (p<0.001) (34). This population corresponds to the hematopoietic stem and progenitor cells in humans (39). These results suggest that the enhanced LUC population in the peripheral blood of mtFVB mice after 5-FU treatment might be due to an aging hematopoietic system. Additionally, compared to mtAKR, a trend to less erythroid cells in the bone marrow of old mtFVB mice and a significantly earlier decrease in B cells were observed. This also points to an aging of the hematopoietic system, although the exact mechanisms have to be elucidated.
In summary, the amino acid exchanging mitochondrial nt7778 G/T polymorphism impacted first and foremost the ROS levels in murine BMC. While ROS levels decreased significantly with age, ATP levels were unaffected. Moreover, significantly lower hemoglobin content in erythrocytes was detectable in older mice. Stress induction by cytostatic administration revealed no homogenous marked differences between both strains investigated.
The underlying signaling cascades leading to the effects of the mtDNA variation remain to be investigated, as well as the different impact on various cell types and tissues concerning ROS levels.
Acknowledgements
The Authors would like to thank the animal attendants of the central animal facility of the University of Rostock for providing the facilities. The Authors thank Emma Woodham for the critical reading of the manuscript and helpful advice.
This work was funded by the German Federal Ministry of Education and Research (BMBF) as part of the GerontoSys initiative (ROSAge project), FKZ 0315892A.
Footnotes
↵* These Authors contributed equally to this work.
This article is freely accessible online.
Conflicts of Interest
There is no conflict of interest to declare.
- Received August 31, 2016.
- Revision received September 15, 2016.
- Accepted September 16, 2016.
- Copyright © 2016 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}