


Abstract
Aim: We hypothesized that radiation-induced fibrosis was, in part, a result of altered signal transduction that directly modulates integrin expression and may indirectly affect the extracellular matrix (ECM). Major focus was given on protein kinase C (pKC). Materials and Methods: Rat FRTL-5 and primary thyroid cells were exposed to proton radiation (5 and 10 Gy). Hours to days after exposure, a series of assays were performed. In addition, the neck region of Lewis rats was proton-irradiated to 40 Gy (5 Gy/day or 10 Gy/day). At 11 weeks after exposure, thyroid tissue was evaluated. Results: Accumulation of ECM in irradiated FRTL-5 and primary thyroid cells was coincident with loss of tissue organization and follicularization at one or more doses and time points. Several pKC isoforms increased post-irradiation, which coincided with modulated integrin expression; fibronectin, laminin and collagen were also altered (p<0.05 vs. 0 Gy). Modulation of thyroid cells in culture with 12-O-tetradecanoylphorbol-13-acetate (TPA)±calphostin C supported a direct role of pKC in these altered properties. Thyroid tissue from irradiated rats had significantly more fibrotic lesions and increases in several pKC isoforms, integrins and fibronectin compared to 0-Gy (p<0.05). Conclusion: pKC is a likely contributor to alteration of key players associated with radiation-induced fibrosis.
Radiation is a major treatment strategy for the control and potential eradication of cancer. Radiotherapy is successful for a majority of patients, although there are detrimental side-effects that continue to be the focus of scientific research in attempts to improve the outcome (1, 2). In particular, when large volumes of tissue are irradiated a significant sub-population of patients has ensuing fibrosis, which ultimately results in morbidity and eventual mortality. There have been many studies of fibrosis, including those on experimentally-induced models that have revealed some of the components that participate in the pathway of uncontrolled wound healing. Nevertheless, there remain unanswered questions regarding many aspects of radiation-induced fibrosis, including the triggers and physical changes that initiate the process. Toward that end, we proposed to apply our understanding over the inflammatory process, which also culminates in fibrotic changes, to that of fibrosis mediated by radiation exposure.
Radiation injury results in inflammatory changes that are strikingly similar to those found at sites of autoimmune disease. From our previous studies using a thyroid model of autoimmune thyroiditis (3-8), as well as published information from other laboratories (9), we know that an initial change that accompanies inflammatory and radiation injury is modulation of the signal transduction pathway leading to an increase in protein kinase C (pKC) expression and enzymatic activity. In autoimmune thyroid disease in rodents (3, 6-8), there is a sustained elevation in pKC activity that is attributable to an increase in the protein corresponding to a number of pKC isoenzymes. If the increase in pKC is sustained following irradiation, it may be similar to what occurs in autoimmune thyroiditis, i.e. uncontrolled phosphorylation alters the function of many integral membrane proteins responsible for maintaining polarity and cell-to-cell cohesion. Indiscriminate phosphorylation is a hallmark of pKC, which can hyper-phosphorylate previously phosphorylated Ser/Thr substrates leading to up- and down-regulation of structural proteins, as well as modulate receptor/ligand-mediated signaling (10).
The structural components of ECM involved in fibrosis are mainly collagen (CN), fibronectin (FN) and laminin (LN), along with integrins that function as ECM receptors. CN is the most abundant protein accounting for 30% of the total protein in the human body (11). CN IV and LN are present in thin layers of basement membrane (BM) enveloping each follicle in normal rat thyroid glands (12). In advanced cases of liver fibrosis, 6-times more ECM is present compared to normal liver, including CN (I, III and IV), FN and LN (13). FN is an important component of the ECM, second only to CN in quantity, consisting of 20 different homologs (14). FN exists as soluble and insoluble isoforms that play important roles in tissue repair (14-16). FN is present in inflammatory cells and its down-regulation at inflammatory sites delays the wound healing process (17). The levels of insoluble or cellular FN also increase following radiation (18). In hepatic fibrosis, serum LN along with type IV CN can be used as markers of the disease (19). It has been shown that LN signaling involves kinase/phosphatase cascades (20) and mitogen-activated protein kinases (MAPK) (21).
The major cell surface receptors responsible for attachment of the above-mentioned components of ECM are the integrins. The integrins are a family of heterodimeric, transmembrane proteins consisting of two subunits, designated α and β. The α chains link cells to the extracellular environment. Integrin α-4 is involved in pathogenesis of chronic inflammatory diseases, such as rheumatoid arthritis, diabetes type1, inflammatory bowel disease and multiple sclerosis. Thus, blocking integrin α-4 has been proven useful to decrease or delay the development of these disorders (22). It has been demonstrated that certain integrins, including α-5 and α-v, are linked to the fibrotic process in renal fibroblasts (23). pKC-mediated events can affect the activity of integrin-mediated signaling; the activation of phosphoinositol 3 (PI-3) kinase by pKC affects the MAPK pathway via effects on Raf (24).
Collectively, this information supports the possibility that pKC has the potential to be an initiating factor in the aberrant events that lead to fibrotic changes in susceptible subjects. To address this and advance our understanding over the fibrotic process, we used the sensitive Lewis rat strain. We have previously used this strain to induce experimental autoimmune thyroiditis (EAT) (6). Overall, this is an excellent model to use for radiation-induced fibrosis due to the strong association between inflammation and pro-fibrotic changes.
Materials and Methods
Animals. Female Lewis rats (n=80) were purchased from Charles River Laboratories (Wilmington, MA, USA) and housed in the Loma Linda University (LLU) Animal Care Facility. Euthanasia was performed by CO2 asphyxiation. All animal care and treatment procedures were in accordance with the current animal welfare act under a protocol approved by the Institutional Animal Care and Use Committee.
Tissue samples/cell cultures. Thyroid glands from Lewis rats were surgically removed and frozen. The fresh-frozen tissue was sliced vertically in ~10 μm sections starting from the outer edge of the lobe proceeding toward the isthmus and stored desiccated at −70°C. The Fischer rat thyroid cells (FRTL-5) were purchased from the American Type Culture Collection (Manassas, VA, USA) and cultured as previously described (25, 26). The cells were allowed to establish into adherent follicular cultures for 5 days with medium changes when necessary.
For primary cell culture, thyroids from non-irradiated Lewis rats were removed and immediately washed in Hank's balanced salt solution (HBSS) (GIBCO, Invitrogen, Carlsbad, CA, USA) containing penicillin, streptomycin and fungizone (50 kg/ml, GIBCO). The tissue was sliced, minced and then incubated for 60 min at 37°C in 5 mg/ml collagenase (Boehringer Mannheim, Indianapolis, IN, USA). The digested thyroid cells were suspended in medium and dispensed into 6-well plates containing coverslips pre-coated with CN I from rat tail and skin. The cells were cultured in Dulbecco's modified Eagle's medium (DMEM):Ham's F-12 medium (GIBCO) (50:50 vol/vol) supplemented with all essential ingredients as done in our previous studies (6). The cells were incubated undisturbed at 37°C in 5% CO2 for 2 days.
Radiation conditions. Proton radiation was used throughout this study because it is a major form of radiation used for therapy at our Institution and its use is increasing worldwide. Since much less information is available on the biological effects of protons on normal tissues compared to photons, e.g. γ-rays and X-rays that have been utilized for many decades, studies, such as this, are important to perform.
External beam proton-irradiations were performed using 250 MeV protons from the Loma Linda University Proton Treatment and Research Facility synchrotron accelerator. Protons were delivered in 0.3−s pulses every 2.2 seconds. The cells were irradiated in flasks, 6-well plates and on 8-well chamber slides. The entrance dose of the Bragg curve was delivered to the cells with the peak outside the cell area. Doses of 5 and 10 Gy were used for cell irradiation in order to maintain consistency with our in vivo irradiation regimens that were based on a fibrosis-related study by Wehrhan et al. in which the head and neck region of rats was irradiated (27). A study by Xavier et al. also demonstrated that single doses of 35 or 45 Gy were pro-fibrotic in rodents (28).
For irradiating rats, the 100-MeV horizontal beam line was used to deliver the protons to the neck region. The rats were lightly anesthetized with isoflurane while in an acrylic holder. All animals were irradiated through a single anterior field to cover the thyroid with the entrance dose. The Bragg peak was outside the rat body to ensure that posterior structures, such as the esophagus and spinal cord, would not be damaged. The dose delivered was 5 Gy/fraction and 10 Gy/fraction given each consecutive day until 40 Gy was achieved. The 40 Gy total dose was selected because it can induce pro-fibrotic changes (27).
Immunocytochemistry. The slides with fresh-frozen thyroid sections or cells were fixed prior to adding primary antibodies as described (7). The primary antibodies were mouse monoclonal pan pKC, mouse monoclonal pKC isoforms α, β, δ, γ, θ, λ and ε (BD Transduction Laboratories, San Jose, CA. USA), polyclonal antibodies pKC ζ and η (Novus Biologicals, Littleton, CO, USA), integrins α-2, -4, -5, -6 (Santa Cruz Biotechnology, Santa Cruz, CA, USA), mouse monoclonal anti-fibronectin (BD Transduction Laboratories), rabbit polyclonal anti-laminin (Novus Biologicals) and pan collagen (ABD Serotec, Oxford, UK). The secondary antibodies were labeled with Alexa Fluor488 dye and the nuclear label was propidium iodide (all from Molecular Probes Inc., Eugene, OR, USA).
For the in vivo study, the time point for analysis (11 weeks post-irradiation) was selected in order to evaluate thyroid status with respect to the process of fibrinogenesis. One half of one lobe of the gland was embedded in paraffin and the other half was cryoprotected; ~10 μm sections were cut and used for immunocytochemistry. The fresh frozen sections were processed as above with the addition of formaldehyde cross-linking to preserve any infiltrating cells.
To quantify specific levels of the immunofluorescently labeled protein, thyroid tissue sections or cells were scanned on the laser scanning cytometer (LSC) with an Olympus BX50 base (Olympus America, Melville, NY, USA) as described previously (26). Briefly, the LSC is configured with argon ion and helium-neon laser for multi-color analysis. Cells were contoured by their red nuclear fluorescence (propidium iodide) and gated by parameters set for signal intensity (fluorescence-labeled proteins). An average of 5,000 cells were scanned per sample using optimized protocol and display settings. Data are presented as means±standard deviation (SD).
Trichrome staining method (Masson). The collagen fibers were stained in tissue using the Masson's trichrome stain kit (Sigma-Aldrich, St. Louis, MO, USA). The instructions of the manufacturer were followed.
pKC activity assay. The cell pellets from FRTL-5 cells were resuspended in 150 μl of pKC extraction buffer (Promega Corp., Madison, WI, USA). The cells were lysed by freeze-thawing and shearing with a 22-gauge needle. The lysates were incubated on ice for 30 min; an aliquot was then taken to determine protein concentration using a Bradford assay. Supernatants were analyzed using a non-radioactive pKC activity kit (Stressgen Bioreagents Corp., Ann Arbor, MI, USA). The assay was performed in a microtiter plate pre-coated with pKC substrate. Samples were added to each well and the reaction was initiated by adding 10 μl of adenosine triphosphate and the plate was incubated at 30°C for 90 min. The reaction was stopped by emptying well contents and addition of phosphospecific substrate antibody at 40 μl/well. The plate was covered with adhesive sealer and incubated at room temperature (RT) for 60 min with intermittent shaking. After incubation, the wells were thoroughly washed with 1× wash buffer. Secondary antibody, anti-rabbit IgG and horseradish peroxidase (HRP) conjugate (1:1,000) were added to each well and incubated at RT for 30 min with gentle shaking every 10 min. The wells were washed thoroughly using wash buffer and 60 μl of the tetramethylbenzidine (TMB) substrate was added to each well and the plate was incubated for approximately 50 min at RT until blue color developed. An acid stop solution was added to each well and the absorbance was measured at 450 nm on a microplate reader (model 680; Bio-Rad Laboratories, Hercules, CA, USA). Relative kinase activity was calculated using values with active pKC.
Modulation of pKC activity. For FRTL-5 cells, 12-O-tetradecanoylphorbol-13-acetate (TPA; Sigma Chemical Corp.) was used to increase pKC activity and calphostin C (Sigma Chemical Corp.) was used to decrease pKC activity. The cultures were incubated with 100 nM TPA at 37°C in 5% CO2/95% air. For pKC activity assay, the cells were lysed in pKC assay buffer as described above and the lysates were stored until immediately before use. For immunohistochemistry, the cells were fixed in 70% chilled ethanol at 2-, 4- and 6-h time points. One sample was left untreated to serve as a control. Calphostin C was added at a concentration of 100 nM/vessel and one sample was left untreated as a control. The tissue was incubated at 37°C in 5% CO2/95% air with constant rocking for 3 and 6 h under lighted conditions (8). The ethanol-fixed cells were used for immunohistochemistry at 0, 3 and 6 h.
Quantification of fibrotic lesions. The fresh frozen sections were stained by the trichome method and images were captured at 40× magnification using the Ellipse program (ViDiTo systems, Kosice, Slovakia). Each image was imported and a point grid was overlayed, which had 40×40 pixel size. The regions in the grid that had blue fibers passing through were counted. All slide labels were concealed so that unbiased counting was maintained. An average value for each set was obtained and plotted on a graph as mean fold change±SD.
Statistical analysis. Statistical analysis of results obtained from protein levels, Western blot analysis and pKC activity assays was performed using one-way analysis of variance (ANOVA) (SigmaStat 2001; SPSS, Inc., Chicago, IL, USA) to compare control and experimental values at each time point after proton irradiation. Proportional difference was judged for significance by chi-square analysis. A p-value of <0.05 was considered significant.
Results
Effect of proton radiation on levels of pKC activity in FRTL-5 cells. The cells were exposed to 5 Gy and 10 Gy proton radiation and lysates were harvested 6 h later. Relative optical density (OD) was used to calculate relative pKC activity per mg of protein. The fold change relative to control is shown in Figure 1a. The activity of pKC increased in irradiated lysates when compared to control lysates. Specifically, lysates of 5-Gy irradiated cells showed a >3-fold increase (p<0.001) and those from 10-Gy irradiated cells showed a >2-fold increase (p<0.05).
Duration of radiation-induced elevation of pKC activity in vitro. pKC activity in irradiated FRTL-5 cells, performed on days 3, 5 and 7 post-exposure to protons, are shown in Figure 1b. There was a ~1.6-fold increase in 10-Gy-irradiated cells compared to control lysates at 3 days, but statistical significance was not obtained. At 5 days post-irradiation, the activity increased nearly by 1.3- and 2.2-fold in 5-Gy and 10-Gy samples, respectively (p<0.001). The pKC activity remained elevated at 7 days with the 5-Gy samples having a ~3-fold higher level than normal (p<0.05). In contrast, the 10-Gy lysates had pKC activity near basal level at 7 days.
Duration of radiation-induced elevation of pKC expression in vitro. FRTL-5 cells from Fischer rats and primary thyroid cells from Lewis rats were exposed to 5-Gy and 10-Gy doses and harvested on days 3, 5 and 7. As shown in Figure 1c, pKC expression on day 3 in primary thyroid cells exposed to 5 Gy was ~2.5-fold greater (p<0.001) and ~1.5-fold greater after 10 Gy than control (p<0.05). Further, increased levels were noted 5 days after radiation with a ~1.7-fold increase in both 5-Gy and 10-Gy samples compared to 0-Gy (p<0.001). At 7 days, pKC expression was still elevated by almost 3.8-fold in 5-Gy samples and by 2.3-fold in 10-Gy samples (p<0.001 vs. 0 Gy). Similarly, Figure 1d shows the duration of radiation-induced elevation in FRTL-5 cells. Expression of pKC was elevated at day 3 by ~2.4-fold in both the 5 Gy and 10 Gy samples (p<0.05 vs. 0 Gy). There were 1.4-fold and 2.6-fold increases after 5 Gy and 10 Gy, respectively, on day 5 (p<0.001 vs. 0 Gy). The elevated pKC protein expression was sustained in FRTL-5 cells so that even 7 days after irradiation it remained higher than controls (p<0.001).

Effect of proton radiation on pKC activity and expression. pKC activity was determined using the pKC kit and levels were measured as optical density read at 450 nm. The relative activity per mg of total protein was calculated and plotted as fold change. To determine the level of pKC protein, immunohistochemical analysis was performed after proton irradiation; results from irradiated cells were compared with the 0 Gy controls at each time point. Panel a: pKC activity in FRTL-5 cells was enhanced approximately 4-fold in 5 Gy samples and 2.5-fold in 10 Gy samples. Panel b: The highest pKC activity in FRTL-5 cells was seen at 7 days after 5 Gy, but was equivalent to normal at 10 Gy. Panel c: In primary thyroid cells from Lewis rats, expression of pKC protein increased in response to radiation and remained elevated up to 7 days. Panel d: Similar results were obtained for FRTL-5 cells with sustained expression of pKC protein for up to 7 days after irradiation. All of the values were recorded as mean fold change±SD from two independent experiments. There were 3 samples analyzed per group. *p<0.05, **p<0.005.
Effect of pKC modulation on thyrocyte expression of integrins. To test whether pKC was responsible for changes in integrin levels, TPA and calphostin C were used to modulate pKC activity in FRTL-5 cells. The basal expression of integrins α-6, α-4 and α-5 are shown in panels a, d and g of Figure 2. The second column (panels b, e and h) shows the levels at 2 h after TPA treatment. Integrin α-6 expression was the most reduced post-TPA treatment compared to non-treated samples. However, integrins α-4 and α-5 increased dramatically when pKC was elevated by TPA. The third column in Figure 2 (panels c, f and i) shows the levels of integrins α-6, α-4 and α-5 in cultures that received TPA for 2 h and then calphostin C for 2 h. The modulation of pKC back to reduced levels revealed that FRTL-5 cells again expressed integrin α-6, but not α-4 and α-5. This is opposite of the findings when pKC was increased using TPA, i.e. the level of integrin α-6 decreased and integrins α-4 and α-5 increased. When the elevated pKC activity was decreased using calphostin C, integrin α-6 returned to its basal state, but integrins α-4 and α-5 were decreased.

The effect of pKC modulation on expression of integrins in FRTL-5 cells. The nine panel composite on the left shows the basal expression of integrins α-6, α-4 and α-5 in the first column (panels a, d and g). The second column (panels b, e and h) shows the levels of integrins α-6, α-4 and α-5 after 2 h treatment with 12-O-tetradecanoylphorbol-13-acetate (TPA). The third column (panels c, f and i) shows the levels of integrins α-6, α-4 and α-5 in cultures that received TPA for 2 h and then calphostin C treatment for 2 h. The experiment reveals that basally the FRTL-5 cells, like most thyroid tissue, expresses integrin α-6 but not α-4 and α-5. However, when pKC levels are increased with TPA, the levels switch with integrin α-6 decreasing and α-4 and α-5 increasing. When the elevated pKC activity was decreased with calphostin C, the levels of all three integrins returned to their basal state. Magnification is 20×. The bar graph on the right shows results based on immunocytochemical analysis of integrins in FRTL-5 cells post-radiation. The cells were grown on 8-well chamber slides, proton-irradiated at 5 Gy and 10 Gy and then harvested 6 h later. Primary antibodies against integrin α-2, -4, -5 and -6 were used. Alexa 488 was used as secondary antibody and propidium iodide as nuclear stain; approximately 5,000 cells were analyzed with a laser scanning cytometer. The values were recorded as mean±SD obtained using two independent experiments. *p<0.05 vs. 0 Gy.
Fold change in expression of pKC isoforms, integrins and adhesion proteins in tissue sections obtained from Lewis rat thyroids.
Measurement of integrin levels after proton irradiation in vitro. FRTL-5 cells were immunolabeled at 6 h post-irradiation with antibodies against integrins α-2, α-4, α-5 and α-6. The levels of expression are shown in the bar graph of Figure 2. Radiation caused a decrease in integrin α-2 compared to control cells, but this was not statistically significant. In the case of integrins α-4 and α-5, there was a significant increase up to 1.6-fold and 4.5-fold, respectively, in cells that received 10 Gy (p<0.05 vs. 0 Gy). Additionally, the expression of integrin α-6 showed a significant decrease of ~70% in 5-Gy irradiated cells (p<0.05 vs. 0 Gy).
Modulation of pKC activity using calphostin C. The FRTL-5 cells were incubated with calphostin C and immunolabeled with pan-pKC and integrin α-4, α-5 and α-6. As shown in Figure 3 top left panel, pKC increased with radiation alone up to ~1.2-fold. Upon calphostin C treatment, at 3 h, the pKC levels dropped by ~30% in both 5-Gy and 10-Gy samples compared to radiation alone (p<0.01). At 6 h, the effect of calphostin C was still apparent with pKC levels decreased ~70% compared to samples that received only radiation (p<0.05).
To demonstrate the effect of pKC inhibition on integrin levels, the cultures were treated with calphostin C and then immunolabeled for integrins α-4, α-5 and α-6. In Figure 3 top right panel, the levels of integrin α-4 are graphed with and without calphostin C and irradiation. In the case of radiation (no calphostin C), the levels of integrin α-4 increased ~1.3-fold post-5-Gy and 2.1-fold post-10-Gy. However, after treatment, the levels dropped in all samples. Calphostin C alone without radiation dropped the levels 20% at 3 h (p<0.05) and 70% after 6 h (p<0.001). In the 5-Gy samples, the drop was ~33% at 3 h and 81% after 6 h of treatment when compared to 5-Gy untreated sample (p<0.001). In the 10-Gy samples, there were decreases of ~74% and 77% after both 3 h and 6 h of calphostin C treatment when compared to untreated 10-Gy samples (p<0.001).
The expression of integrin α-5 increased ~1.6-fold with 5-Gy and ~1.7-fold with 10 Gy radiation, as shown in Figure 3 bottom left panel. After treatment with calphostin C, integrin α-5 levels decreased 4% after 3 h and 50% after 6 h (p<0.001) in non-irradiated controls. In the 5 Gy samples, the levels decreased ~38% after 3 h and 60% after 6 h of calphostin C treatment compared to untreated samples (p<0.001). With the higher 10 Gy dose, the elevated levels decreased to ~40% at 3 h and 60% at 6 h as compared to untreated irradiated samples (p<0.001).
To determine whether there was a relationship between decreased levels of integrin α-6 and enhanced pKC activity, we modulated the pKC levels. In the TPA-induced enhancement of pKC activity we observed a decrease in integrin α-6. Hence, we decided to elevate pKC, using radiation, and then inhibit the enhanced pKC using calphostin C and measure integrin α-6 levels. As shown in Figure 3 bottom right panel, integrin α-6 decreased with radiation alone. However, when the cells were treated with calphostin C, integrin α-6 increased. After 3 h, the calphostin C-treated samples increased ~6-fold in 5 Gy and ~4-fold in 10-Gy samples (p<0.001 vs. 0 Gy non-treated samples).
Induction of fibrosis in vivo. The rats were divided into 3 groups: control, 5 Gy/day (8 times to achieve 40 Gy total) and 10 Gy/day (4 times to achieve 40 Gy). General health was monitored by weighing them every 2 days. An initial effect of radiation was hair loss in the area where the beam exited the neck. Although there was some weight loss in the first few days, the rodents quickly regained normal weight and remained healthy thereafter. A fully established fibrotic thyroid was observed at 11 weeks after irradiation.
Thyroid tissue sections were stained for collagen, since it is a main component of fibrotic lesions. As shown in Figure 4, the collagenous structures stained bright blue. Panel a shows a representative section of non-irradiated rat thyroid with little blue surrounding the cells and intact tissue organization within the follicle. However, in sections from irradiated rats (Figure 4, panels b and c), there was an increase in filamentous structures, which were mainly composed of collagenous fibers and spanned across the tissue sections. To further confirm our results, fibrotic regions were scored blindly, i.e. objectively. Approximately 10 images were scored per rat after 0 Gy, 5 Gy fractions and 10 Gy fractions. Mean fold change±SD (n=40) is shown in Figure 4d. There was a ~2-fold increase in fibrotic lesions in irradiated rat thyroids (5 Gy and 10 Gy fractions) (p<0.05 vs. 0 Gy).

Calphostin C treatment of FRTL-5 cells. Calphostin C (Cal C) is an inhibitor of pKC activity. The top left panel shows the pan pKC expression±radiation and±Cal C. The pKC expression increases post-proton irradiation. Cal C was used to decrease the radiation-enhanced pKC at 3 h and 6 h. The top right panel shows a decrease in integrin α-4 levels with dose. The levels came back up with Cal C treatment suggesting possible role of pKC in modulating integrin α-4 levels. The bottom left panel shows induction of integrin α-5 in irradiated cells as compared to unirradiated. The radiation-induced integrin α-5 was reduced using Cal C. The bottom right panel depicts the increase in integrin α-6 expression post-Cal C treatment. Note that the y-axis for integrin α-6 is higher than the rest. Values are means±SD. *p<0.05, **p<0.001 vs. No treatment (Tx).
Immunohistochemical measurement of pKC isoforms. After inducing fibrosis in irradiated Lewis rats, levels of pKC isoforms were obtained 11 weeks post-exposure to protons (Table I). When 5 Gy/fraction was used, there were significant increases in pKC α, ε, θ and η (p<0.001 vs. 0 Gy), as well as PKC γ and ζ (p<0.05 vs. 0 Gy). After 10 Gy/fraction, high levels were noted for PKC α and η (p<0.001 vs. 0 Gy) and for PKC θ and ζ (p<0.05 vs. 0 Gy). The level of pKC γ expression increased ~2.6-fold in 5-Gy and 1.5-fold in 10-Gy fractionated samples. Additionally, pKC ε, η and ζ increased nearly 2-fold, 7-fold and 4-fold, respectively, in fibrotic thyroids compared to controls. Representative images of increased expression of pKC γ (panels a-c) and pKC ζ (panels d-f) are shown in Figure 5.
Integrin levels after radiation-induced fibrosis. The levels of several integrins are summarized in Table I. In irradiated thyroids, integrins α-2, α-4 and α-5 increased 1.3-fold (10-Gy fractions), 5-fold (5 Gy fractions, p<0.001) and 3-fold (5-Gy fractions; p<0.001) compared to non-irradiated controls. There was approximately a 40-50% decrease in integrin α-6 in irradiated samples versus controls (p<0.05). The images of integrins α-4 and α-6 are shown in Figure 5 panels g-j. As the thyroid is a laminar matrix, there were high levels of integrin α-6 and nearly undetectable levels of integrin α-4 in non-irradiated thyroids. A significant decrease in integrin α-6 expression and a significant induction in integrin α-4 expression were recorded in thyroid sections of the rats that received 10 Gy/fraction.

Trichrome staining of Lewis rat thyroid tissue 11 weeks after irradiation. The thyroid sections obtained from proton irradiated (5 Gy and 10 Gy fractions; total=40 Gy) and control rats were stained using a trichrome kit, which stains collagen fibers blue, nuclei blue/black and intercellular fibers red. Panel a depicts the normal structures in a representative section of unirradiated rat thyroid. Panels b and c clearly show that radiation increased collagenous structures that stained blue spanning across the tissue section. The images were imported in Ellipse software. Each image was overlapped with a PointGrid type of stereology and size of the grid was chosen to be 40×40 pixels. The regions within each grid that contained any blue staining were counted using Ellipse software as described in the Materials and Methods section. The images are at 40× magnification and the fold change is graphed (mean±SD) in panel d. *p<0.05 vs. 0 Gy.
ECM components after proton radiation-induced fibrosis. As shown in Table I, there was a ~1.3-fold increase in fibronectin in irradiated thyroid sections (10 Gy fractions) when compared to controls (p<0.001). Also, in the case of pan collagen, significant increases of 3-fold and 2.6-fold were recorded in irradiated fibrotic tissues receiving 40 Gy by 5 and 10 Gy fractions compared to controls (p<0.001). Decreases of ~30% (5 Gy fractions; p<0.05) and 20% (10 Gy fractions) were measured in laminin levels in irradiated tissue.
Discussion
The premise of this study was that thyroid epithelial cells are functionally dependent on their integration into an organized glandular assembly. The hierarchy of macromolecule and protein interactions proceeds from the ECM and integrins, to tight and adhesion junctions, which in turn provide the proximity required for gap junctions to assemble in plasma membranes of the contacting cells. We proposed that hyper-phosphorylation of structural proteins and ECM receptors by radiation-induced increases in pKC would alter the physical cohesion of thyrocytes leading to an accumulation of fibronectin and collagen, thus ultimately contributing to fibrosis. Previous work in our laboratory with an autoimmune thyroiditis model suggested that there were changes in normal thyroid organization that may facilitate progression from chronic inflammation to fibrosis (3-8). In the present study, we observed a radiation-induced loss of integrin α-6 (laminin receptor) in thyrocytes (Table I). Since the thyroid is a laminar matrix, high levels of the laminin receptor and negligible levels of fibronectin receptors (integrins α-4 and α-5) were found in normal tissue, as expected. However, similar levels were induced in the inflamed thyroids in our previous study. These data suggested that there may be correlation between the enhanced pKC levels and change in integrin expression.
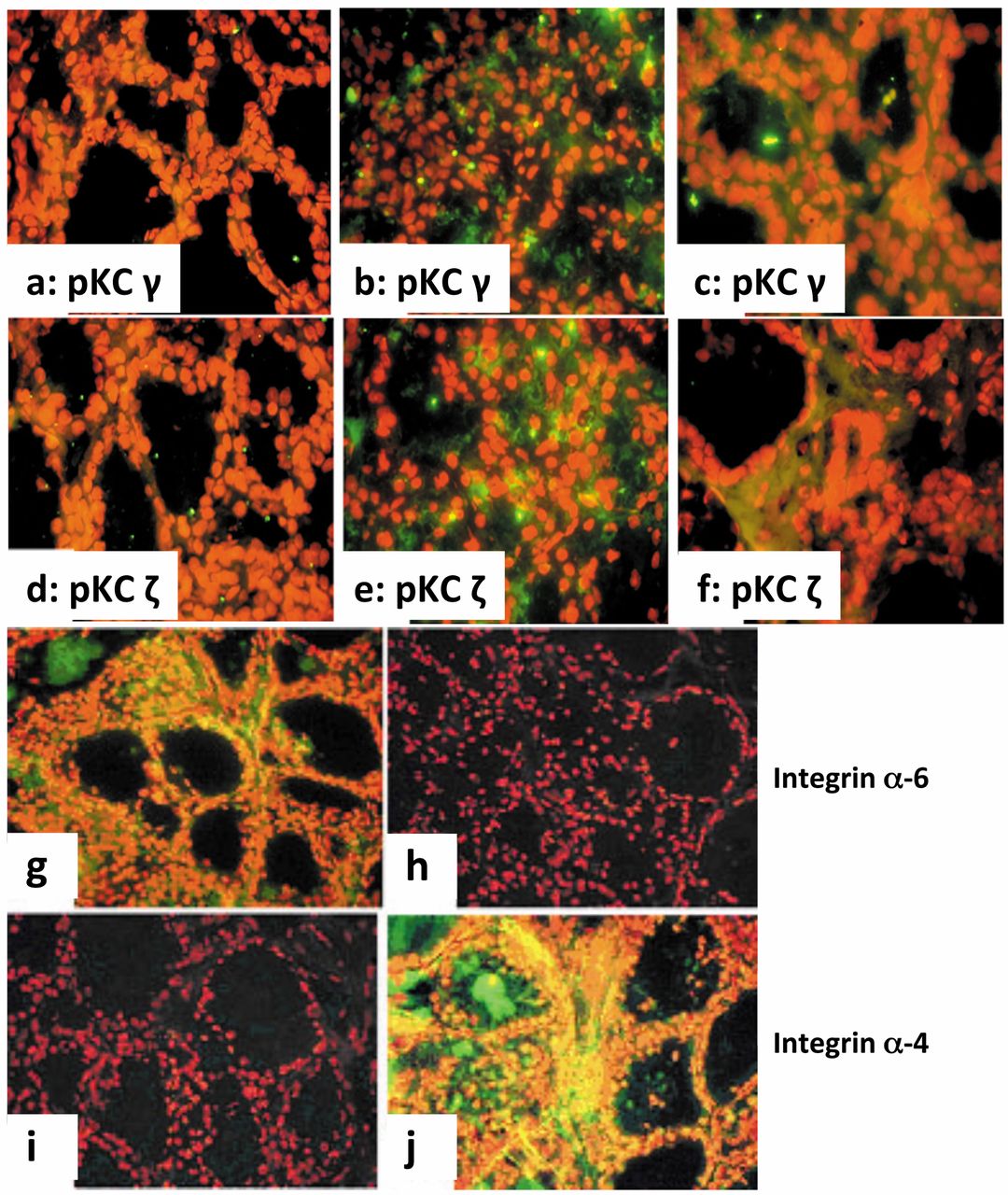
Immunohistochemical detection of pKC isoforms and integrins in Lewis rat thyroid glands 11 weeks after radiation-induced fibrosis. Thyroid sections from proton-irradiated and non-irradiated rats were harvested post-radiation and immunolabeled as described in the Materials and Methods. Panels a and d show levels of pKC γ and pKC ζ in non-irradiated tissue, respectively; panels b and e show expression after 40 Gy irradiation in 5 Gy fractions; panels c and f show expression after 40 Gy irradiation in 10 Gy fractions. Both radiation regimens resulted in increased expression of these two pKC isoforms. Representative sections stained for integrins α-6 and α-4 on thyroid epithelium are shown in the bottom four panels. Panels g and i from non-irradiated rats show that integrin α-6 is expressed, but integrin α-4 is not expressed. However, the irradiated tissue was devoid of integrin α-6 (panel h) and integrin α-4 was induced (panel j). Magnification used is 60×.
TPA is a highly potent tumor producer in vivo and activator of pKC in vitro. TPA increased pKC activity in our mouse and rat studies between 40-60% above basal levels, which is similar to the ~40% increase in the thyroiditis model (3, 6, 8, 26). We modulated pKC once again using TPA in non-irradiated cells and measured alterations in integrins α-4, α-5 and α-6. Normal thyroid cells treated with TPA had high pKC expression and activity; there was also an increase in integrins α-4 and α-5. The expression of integrin α-6 decreased in the TPA-treated cells compared to untreated controls. This experiment established the fact that increases in pKC were, in part, responsible for modulating integrins in the rat thyroid cells.
Since radiation injury results in inflammatory changes that are strikingly similar to those found at sites of autoimmune disease and activation of pKC is one of the early responses to ionizing radiation (29, 30), we sought to determine whether radiation induced similar pKC enhancement in our thyroid model. Preliminary experiments with FRTL-5 cells (normal thyroid cell line) exposed to γ-radiation indicated that several pKC isoforms increased as early as 3 h after irradiation.
This led us to question whether FRTL-5 cells exposed to proton radiation would give similar results. In contrast to photons (γ-rays, X-rays), proton radiation can be targeted more precisely, thus minimizing side-effects, such as burns, at the entry site. Although damage to normal tissue surrounding a tumor cannot be negated, many medical centers are opting for protons as an alternate to conventional radiotherapy. Since the Loma Linda proton accelerator is readily available for research, it was logical to use proton radiation for our studies. FRTL-5 cells were exposed to proton radiation and the overall expression of pKC protein was determined using a pan-pKC antibody. There was an increase in pKC expression, as well as activity, in the irradiated cells compared to controls. Specifically, maximum activity was recorded at 5-Gy protons, thereby suggesting that there was increased pKC phosphorylation in the samples irradiated in vitro. Hence, for FRTL-5 cells there was a dose-dependent increase in pKC with 5 Gy being the threshold dose beyond which the level did not increase.
In inflammation, pKC activity is sustained in culture long after the infiltrating lymphocytes have been removed. This suggests that there may be inherent modification or autocrine activation that maintains the elevated levels of pKC and the kinase activity. Since this possibility had never been addressed following irradiation, FRTL-5 and primary thyroid cells were used to answer this question. The cells were irradiated at 5 Gy and 10 Gy and harvested at 3, 5 and 7 days. There was increased overall pKC expression in irradiated cells and a high level was sustained even by day 7. In primary thyroid cells, the maximum levels were observed in 5 Gy-irradiated cells on day 7. Whereas, in FRTL-5 cells, maximum pKC expression was recorded in 10 Gy-irradiated cells at 5 days, but it was still higher than controls on day 7.
The results obtained from quantitative immunohistochemistry were further corroborated by pKC activity assays using FRTL-5 and primary cell lysates. It was observed that pKC activity in irradiated lysates was higher on days 3 and 5 than in the non-irradiated samples. Maximum activity was observed at 5 Gy on day 7, which was similar to that observed with immunohistochemistry. It should be noted that the activity in 10-Gy irradiated lysates was approximately equivalent to the basal activity at 7 days, thereby suggesting that a threshold was reached beyond which the activity did not increase irrespective of radiation dose or incubation time. However, the possibility exists that the relatively low pKC activity, as well as pKC level (described above), on day 7 after 10 Gy may be at least partly related to cell death.
In the autoimmune thyroiditis model, it was observed that even though the follicles within diseased thyroids appeared intact, there was loss of coupling between the thyrocytes (unpublished data). Also, it was reported that with the enhanced levels of pKC, there was an induction of integrin α-4 and integrin α-5 (fibronectin receptors) and a reduction in integrin α-6 levels (laminin receptor). The levels of integrins after TPA treatment also showed similar alteration with the artificial enhancement of pKC. However, the effect of radiation on integrins in vitro had not been investigated extensively before. The presented data suggest that there is a significant increase in integrin α-4 and α-5 levels following radiation, as well as a decrease in integrin α-6. These data confirmed our findings from the inflammation study suggesting that inflammation and radiation generate similar responses in thyroid cells.
In earlier experiments, we established that an increase in pKC expression and activity resulted in modulation of integrin levels in vitro. Hence, to establish whether inhibition of pKC resulted in the return of integrins to normal levels, irradiated cells were treated with calphostin C, a potent inhibitor of pKC phosphorylation. The presented data suggest that enhancement of pKC following radiation modulated levels of integrins and inhibition of radiation-induced pKC resulted in reversal of this effect. Thus, pKC may be partly responsible for the “switch” between fibronectin receptors and the laminin receptor. This, in turn, implies that there could be an increased attraction of fibronectin at the site of injury and a reciprocal loss of laminin, which results in development of fibrosis in irradiated tissue and loss of normal glandular function.
An in vivo model of radiation-induced fibrosis was developed using Lewis rats, a strain that is highly susceptible to inflammation. The 40 Gy used to irradiate the rat thyroid gland is comparable to humans receiving 70 Gy radiotherapy of the head and heck as calculated by the Time Dose Fractionation Model or 80 Gy by the Linear Quadratic Model (27). The design of this investigation was modified from a study by Wehrhan et al., wherein they irradiated male Wistar rats with 10 Gy fractions for a total dose of 40 Gy of X-rays (27). Our samples were harvested 11 weeks following proton irradiation. Since, we were interested in side-effects of radiation on normal thyroid tissue as a possible consequence of radiation therapy, we delivered protons at the entrance region to the thyroids, with the Bragg peak falling outside of the animal. No overt signs of sickness were noted during the entire 11-week follow-up after irradiation; hair and weight loss shortly after irradiation were transient.
There was an extensive loss of follicular arrangement in fibrotic tissue. It was interesting to note that the severity of tissue disruption was greater with 5 Gy than with 10 Gy fractionation. Since fibrotic lesions are mainly composed of collagen fibers, thyroid sections were stained so that they could be visualized. As expected, the irradiated fibrotic thyroids had approximately 2-fold increase in collagen fibers compared to controls. It was interesting to note that there was no significant difference between 5 Gy and 10 Gy fractions.
The levels of pKC and integrins were also documented in fibrotic thyroids from irradiated rats to determine whether changes were similar to those recorded in inflammation and in vitro experiments. An important aspect was to determine if the increase in pKC activity induced by radiation was maintained in vivo. We had evidence that this scenario was true in inflammatory diseases like autoimmune thyroiditis, but did not know whether this would occur following radiation exposure. The fibrotic thyroids showed increased expression of pKC isoforms compared to controls, thus confirmed our hypothesis. The up-regulation of pKC isoforms, in turn, modulated integrin expression, with integrin α-6 being down-regulated and integrins α-4 and α-5 up-regulated. The induced expression of the fibronectin receptor (α-4) indicated that the thyroid tissue had accumulated fibronectin bound to the thyrocyte surface. Additionally, although there was accumulation of collagen, there was no significant increase in the classical collagen receptor, integrin α-2. Hence, it may be that the exaggerated collagen accumulation that was seen at fibrotic sites was derived from myofibroblasts. Thyrocytes do elaborate their own collagen that could be regulated in a biofeedback loop with its receptor expression. Additionally, it could also mean that integrin α-2 is not receiving altered signals and, thus, is not involved directly in the collagen-related fibrotic changes.
In conclusion, the accumulation of ECM in thyroid tissue was coincident with a loss of tissue organization and follicularization. This study provided insight into the fibrotic changes associated with radiation injury. Evaluation of the thyroid at time points earlier than the 11 weeks post-irradiation performed in the present study, should be done to follow evolution of the response that leads to fibrosis and to determine if the changes reported here are similar to those recorded for inflammatory injury. Overall, the results suggest that pKC is, at least partly, responsible for the alterations observed in radiation-induced fibrosis. Increase in pKC is responsible for induction of integrins α-4 and α-5 and reduction of integrin α-6. In vitro modulation of thyroid cultures supported the direct role of pKC in these altered properties. The levels of fibronectin and collagen increased significantly post-irradiation in fibrotic thyroids from Lewis rats and there was a decrease in laminin. The increase in collagen was not associated with receptor integrin α-2. In the future, it would be interesting to modulate pKC in vivo and identify the mechanisms by which it alters expression of the integrins and clarify the source of collagen accumulation. The mechanisms by which inflammation and radiation enhance pKC activity and expression should also be investigated.
Acknowledgements
This study was supported by internal funds supplied by Dr. James M. Slater, M.D., F.A.C.R. and the Department of Radiation Medicine at the Loma Linda University Medical Center. The Authors would like to thank Leticia S. Ortloff for her support in the laboratory and Steven Rightnar for assistance with proton irradiation.
- Received December 18, 2014.
- Revision received January 18, 2015.
- Accepted January 20, 2015.
- Copyright © 2015 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.