


Abstract
Background: The purpose of the present study was to compare the antioxidant potential of lipophilic tea polyphenols (LTP) against the one of naturally-occurring water-soluble green tea polyphenols (GTP) in a two-stage model of diethylnitrosamine (DEN)/phenobarbital (PB)-induced hepatocarcinogenesis in Sprague-Dawley rats. Materials and Methods: GTP/LTP was given 5-times weekly by oral gavage with tea polyphenols equivalent to 0-, 40- and 400-mg/kg of body weight/day. GTP/LTP treatment was started 2 weeks prior to the initiation of DEN and continued for 30 weeks. Results: Histopathological and electron microscopic examination of liver tissue confirmed the protective effect of LTP on DEN/PB-induced liver damage and pre-carcinogenesis. LTP treatment significantly increased total antioxidant capacity (T-AOC) and glutathione peroxidase (GSH-Px) activity in liver tissues. Immunohistochemical detection of cellular nuclear factor erythroid-2-related factor-2 (Nrf2) and peroxiredoxin-6 (P6) indicated a down-regulation in Nrf2 and up-regulation of P6 expression in the liver of LTP-supplemented rats. Conclusion: The present study provides evidence for the first time, that LTP exerts significant antioxidant effects on DEN/PB-induced liver damage and hepatocarcinogenesis through elevating T-AOC levels, enhancing GSH-Px activity and inducing P6 expression in rat liver tissues.
Hepatocellular carcinoma (HCC) is the sixth most common cancer and the third leading cause of cancer-related mortality worldwide (1). It is well-reported that the major risk factors for HCC are chronic liver inflammation due to infection by the hepatitis B virus (HBV) and/or hepatitis C viruses (HCV), long-term alcohol consumption and exposure to carcinogenic toxins (2). Unfortunately, HCC is frequently diagnosed at late stages because of absence of clear pathognomonic symptoms. As a result, several patients have untreatable HCC by the time they are first diagnosed (3). Continuous oxidative stress, resulting from generation of reactive oxygen species (ROS) by environmental factors or cellular mitochondrial dysfunction, may play an important role in hepatocarcinogenesis (4-6). Diethylnitrosamine (DEN), one of the most important environmental carcinogens, may cause the generation of ROS, resulting in oxidative stress and cellular injury. This may account for the carcinogenic effects of DEN (7). Phenobarbital (PB) is not genotoxic, but a two-year feeding study found a dose-related increase in pre-neoplastic hepatocellular lesions (8). PB strongly and rapidly induces liver tumors in animals, when it is administered together with an environmental carcinogen, such as DEN (9).
In view of the limited treatment options and the grave prognosis of HCC patients, preventive measures have been emphasized in dealing with this disease. Since oxidative stress has been implicated in the multi-stage hepatocarcinogenesis process (7, 10), bioactive compounds having antioxidant potentials could be capable of reducing DEN/PB-induced hepatic damage and hepatocarcinogenesis. Green tea polyphenols (GTP) are dietary agents that may hold promise in the prevention of human cancers through its antioxidant activity (11). GTP, a mixture of biologically-active polyphenols in green tea, is mainly comprised of (−)-epigallocatechin-3-gallate (EGCG), (−)-epigallocatechin (EGC), (−)-epicatechin-3-gallate (ECG), and (−)-epicatechin (EC), among which EGCG is the most abundant constituent (12). Blood levels of EGCG are generally lower than 0.5 μM in animals receiving tea preparations in cancer prevention studies due to the compound's limited bioavailability. This is much lower than the concentrations used in in vitro studies of the effects of GTPs (13). The limited bioavailability of natural GTP may reduce its beneficial effects in vivo. These effects may include antioxidant, anti-cancer, anti-obesity, anti-atherosclerotic, anti-diabetic, anti-viral, anti-bacterial, and anti-fungal effects. GTPs may also exert neuroprotective effects (13-18). Lipophilic tea polyphenols (LTP) are GTP compounds chemically modified by an acylation reaction. This modification improves lipid solubility by increasing the number of fatty-acid groups. The biological activity of these compounds can be stabilized, and their bioavailability is significantly increased in comparison to GTPs (19, 20). The antioxidant efficacy of LTP may be increased over that of GTPs, due to its enhanced cellular absorption in vivo. Thus, we infer that LTP has better antioxidant potential than GTPs in vivo which make it possible to reduce DEN/PB-induced hepatic damage and hepatocarcinogenesis.
Nuclear factor-erythroid 2-related factor-2 (Nrf2) is commonly recognized as a redox-sensitive transcription factor that controls the expression of several antioxidant enzymes. These enzymes may protect cells against oxidative stress from a variety of physiological and environmental stimuli (21). Nrf2 responds to oxidative stress by binding to the antioxidant response element (ARE) in the promoter of genes coding for antioxidant and detoxicating enzymes such as NADPH: quinone oxidoreductase 1 (NQO1) and proteins for glutathione synthesis (22, 23). Peroxiredoxin 6 (P6) is one of the ARE-responsive genes regulated by Nrf2. There is a cis-acting element termed ARE in the promoter of P6 gene, and ARE within the P6 promoter is a key regulator of basal transcription of the P6 gene (24). P6 acts as a bi-functional enzyme possessing peroxidase activity and phospholipase A2 activity. This means that P6 may play important roles in antioxidant defense based on its ability to reduce peroxidized membrane phospholipids and in phospholipid homeostasis based on its ability to generate lysophospholipid substrate for the re-modeling pathway of phospholipid synthesis (25). The regulation of Nrf2/P6 by tea polyphenols could strengthen the body's antioxidant defenses, providing chemoprevention and cytoprotection against DEN/PB-induced hepatocarcinogenesis in rats.
GTP-mediated antioxidant activity has been demonstrated in a wide variety of animal models, in which ROS accumulation causes a broad range of diseases and/or pathological changes (26-28). The antioxidant potentials of LTP have not been thoroughly explored in vivo. In the studies reported here, we tested the antioxidant effect of LTP compared to GTP in a rat model of chemically-induced hepatocarcinogenesis. We tested the hypothesis that LTP will be more effective than GTP at inhibiting DEN/PB-induced hepatic damage and hepatocarcinogenesis due to the increased bioavailability of LTP. To the best of our knowledge, this is the first in vivo study to demonstrate the antioxidant effect of GTP and LTP against chemically-induced hepatocarcinogenesis.
Materials and Methods
Animals and diet. Pathogen-free male Sprague-Dawley rats, initially weighing 140-155 g, obtained from the Zhejiang Experimental Animal Center, were maintained at a conventional animal facility in the Laboratory Animal Center of Zhejiang University, China. The animals were housed in automatically controlled conditions with a 12-h light-dark cycle, at 23-25°C room temperature and 50-60% relative humidity. All rats were given standard rodent pellet food and water ad libitum. The experimental protocol was approved by the Laboratory Animal Center of Zhejiang University, and strictly adhered to during the entire study.
Hepatocarcinogenesis model. Experimental hepatocarcinogenesis was initiated by diethylnitrosamine (DEN) and promoted by phenobarbital (PB). DEN dissolved in saline was injected once, intra-peritoneally (i.p.), at a dose of 150 mg/kg body weight. Beginning from the day of the DEN injection, the PB promoter was incorporated into the animals' drinking water at the concentration of 0.05% (w/v).
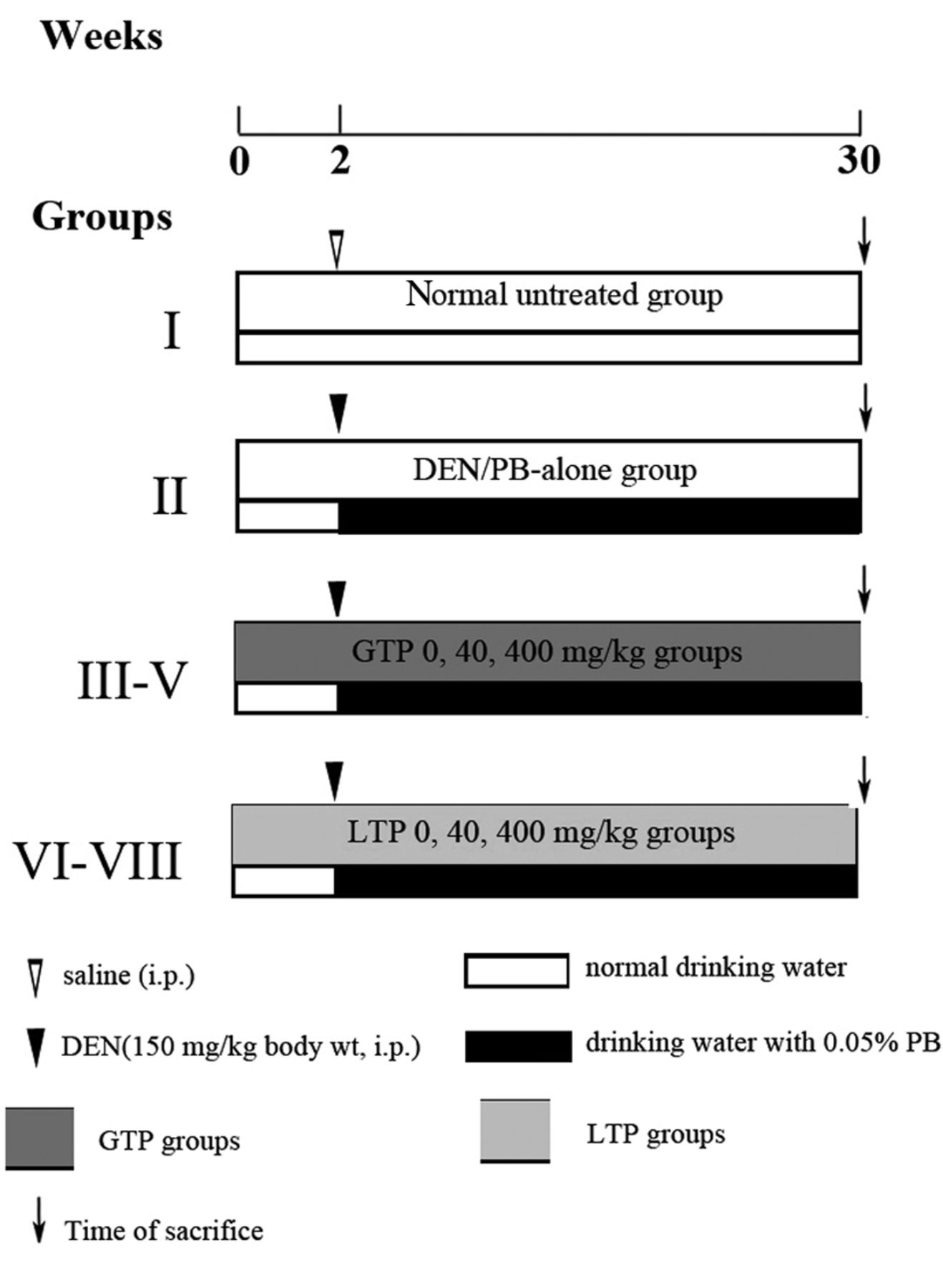
Experimental design and treatment. Rats were randomly divided to 8 groups with 25 animals each: group I (normal untreated group), group II (model group, or DEN/PB-alone group), group III-V (GTP 0-, 40-, 400- mg/kg groups), group VI-VIII (LTP 0-, 40-, 400-mg/kg groups) (Figure 1). At week 2 Group I was administered normal saline intraperitoneally once, meanwhile the other groups were given DEN/PB to stimulate hepatocarcinogenesis. Before the administration of the carcinogen, rats were pre-treated with GTP and LTP for 2 weeks. GTP groups were given GTP at a dose of 0-, 40-, 400-mg/kg of body weight 5-times weekly by oral gavage for 30 weeks, sterile water was given to the 0- mg/kg group. LTP groups were given LTP at a dose of 0-, 40-, 400- mg/kg body weight 5-times weekly by oral gavage for the same period, corn oil was given to the 0-mg/kg group.
Weighing and sample preparation. Body weight was measured once a week, and food consumption was measured once a month. Necropsy was performed immediately after bleeding the femoral artery to death under anesthesia at the end point after starvation for 16 h. Livers were weighed and stored at −80°C for biochemical analyses. Representative liver slices were taken immediately immersed in 4% paraformaldehyde and stored at 4°C for histological and immunohistochemical analyses.
Determination of antioxidant activity. Liver samples were homogenized for 5 minutes in saline (1:9 w/v). After centrifugation at 4,000 ×g for 10 min at 4°C, supernatants were used to analyze the levels of MDA, T-AOC and the activity of GSH-Px. The level of MDA, T-AOC and the activity of GSH-Px in liver tissues were measured using appropriate kits (all from Nanjing Jiancheng Bioengineering Institute, Nanjing, China). The T-AOC was a representative of enzyme and non-enzyme antioxidant in the liver. These antioxidants reduced the ferric ion (Fe3+) to ferrous ion (Fe2+). The latter combined with phenanthroline and produced a stable chelate, which could be measured by spectrophotography at a wavelength of 520 nm.

Schematic representation of experimental design employed in the present study.
Immunohistochemical analysis of Nrf2 and Peroxiredoxin 6. After de-paraffinization, target retrieval by a hot water bath (95°C) in citrate buffer and incubation in 0.3% H2O2 and normal goat serum, sections were subjected to immunohistochemistry with a rabbit polyclonal anti-rat Nrf2 antibody (1:200, 4°C, overnight) (Sigma-Aldrich, St. Louis, MO, USA) and a rabbit polyclonal anti-P6 antibody (1:10,000, 4°C, overnight) (Abcam, Cambridge, MA, USA). All specimens were lightly counter-stained with hematoxylin. The numbers of Nrf2- and P6-positive cells per 103 cells were measured with the use of Image Pro Plus.
Statistical analysis. Unless otherwise specified, all data are presented as Means±Standard Error of the mean (S.E.) and significance of the differences between mean values was determined by one-way analysis of variance (ANOVA) using a commercial software program SPSS (Statistical Product and Service Solutions, SPSS Inc, Chicago, IL, USA) except for the body weight data. The body weight data were analyzed by general linear model/repeated measures. A probability level less than 0.05 was used as a criterion for significance.
Results
General observations. None of the rats in the normal group and the model group died during the observational period, whereas some rats in other experimental groups died due to gavage misoperation. During the entire study period, no differences in food intake were noted among the various experimental groups. No hepatic nodules or tumors were visible by naked eye in the liver of all groups.
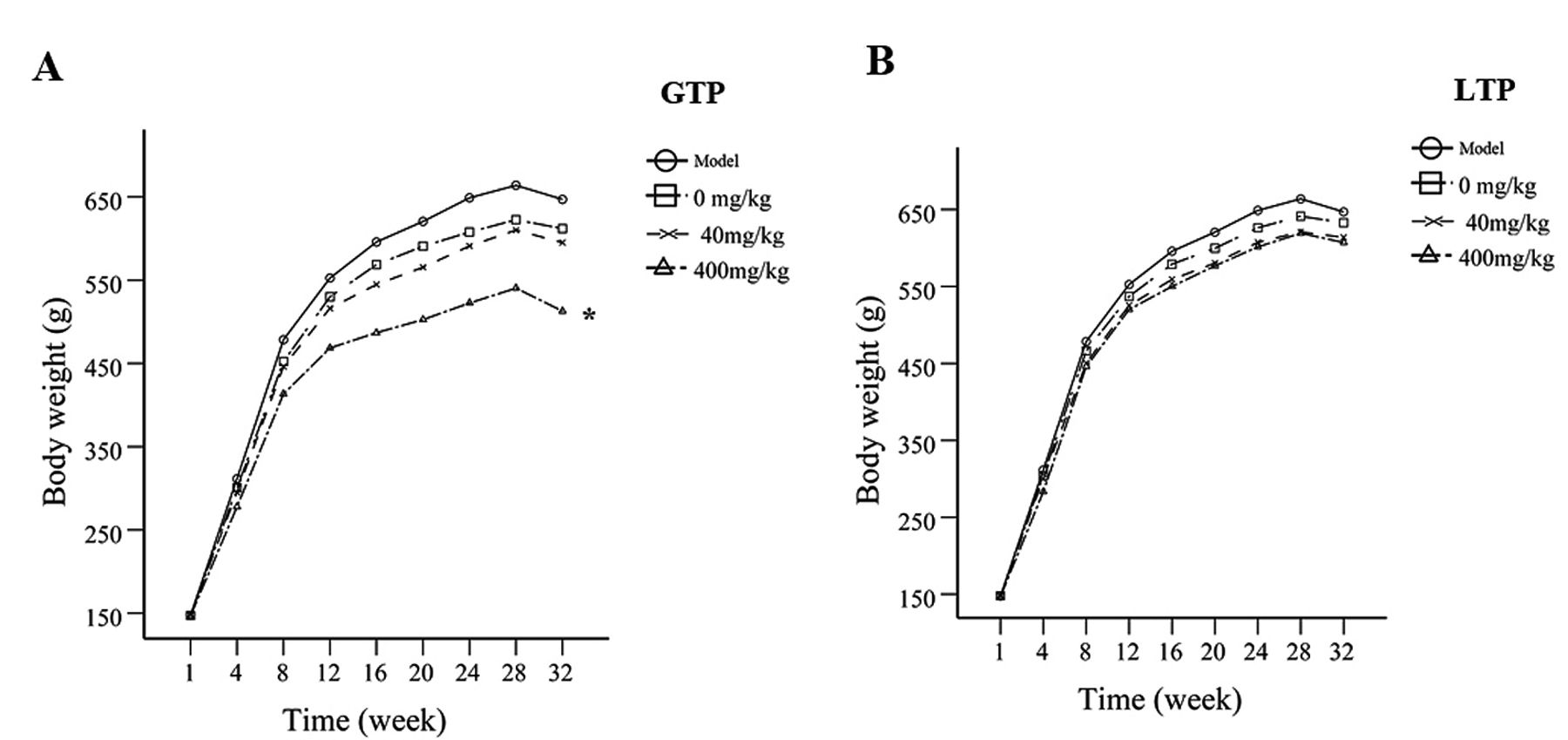
Body weight. Body weight increased in all experimental groups, from an average of 366.2 g to an average of 499.4 g after 30 weeks. The body weight gain of GTP 400-mg/kg group decreased notably (p<0.05) when compared to controls (Figure 2A), while LTP treatments don't show differences within the 3-dose groups (Figure 2B).
Liver and relative liver weight. GTP at 40 mg/kg and 400 mg/kg significantly reduced average liver weight compared with control group (p<0.05). LTP at 40 mg/kg did not alter liver weight but at 400 mg/kg significantly reduced liver weight (p<0.05) (Figure 3A). Significant decreases in relative liver weight of rats in GTP 40-mg/kg group and LTP 400-mg/kg group were noted (p<0.05) (Figure 3B).
Effects of GTP/LTP on hepatic histology. Liver tissue of the normal group revealed normal parenchymal cells with granulated cytoplasm and small uniform nuclei radially-arranged around the central vein. Only animals in the DEN-treated group (model group and 0-mg/kg groups) showed a significant loss of hepatocyte architecture as seen by the presence of extensive vacuolation in the cytoplasm with masses of acidophilic or eosinophilic material. Additionally, the 0-mg/kg LTP group revealed numerous lipid vacuoles. LTP treatment led to gross and noticeable improvement in hepatocellular architecture at the dose of 40 and 400 mg/kg in a dose-response manner (Figure 4A). Electron microscopy shows that exposure to DEN/PB produced an increase of vacuoles in cytoplasm, as well as damages to the mitochondria. The GTP treatment cannot reverse theses damages, while LTP at a dose of 400 mg/kg can moderately reverse these damages (Figure 4B). Microphotograph from LTP 400-mg/kg group is almost similar to that of the normal group. Histopathological and electron microscopic examination of liver tissue confirmed the protective effect of LTP.
Effects of GTP/LTP on antioxidant defense system. LTP at 40 mg/kg and 400 mg/kg significantly increased (p<0.05) total antioxidant capacity (T-AOC) and the same trends were observed in glutathione peroxidase (GSH-Px) activity in liver tissues when compared to the 0-mg/kg group. Whereas GTP only at a high dose of 400 mg/kg can significantly (p<0.05) increase T-AOC and GSH-Px activity (Figure 5).

Animal growth during the entire term of the study. (A) Effect of GTP on body weight gain during DEN-induced hepatocarcinogenesis in rats. Statistical significance was determined by general linear model/repeated measures. *p<0.05 compared to GTP 0-mg/kg group. (B) Effect of LTP on body weight gain during DEN-induced hepatocarcinogenesis in rats.

Effects of GTP and LTP on liver and relative liver weight during DEN-induced hepatocarcinogenesis in rats. (A) Liver weight. (B) Relative liver weight. Statistical significance was determined by ANOVA. *p<0.05 compared to GTP 0 mg/kg group; #p<0.05 compared to LTP 0 mg/kg group. Data are shown as the mean±SE.
Effects of GTP/LTP on antioxidant protein Nrf2 and P6 expression. Immunohistochemical detection of cellular antioxidant protein nuclear factor-erythroid 2-related factor-2 (Nrf2) indicated that Nrf2-positive cells increased significantly (p<0.05) in the livers after exposure to GTP 40 mg/kg, whereas there were few Nrf2-positive cells after exposure to LTP 40 and 400 mg/kg. Many of the immuno-positive cells for Nrf2 were observed in the nucleus, indicating activation of Nrf2 and its subsequent nuclear translocation (Figure 6A, C).

Effect of GTP/LTP on hepatic histology. (A) Histopathological analysis of representative liver tissue from various experimental animals as observed by H&E staining (magnification:×400). (B) Electron Microscope examination of rat liver tissue at the end of the study (magnification: ×8,900).
Immunohistochemical analysis of cellular antioxidant protein peroxiredoxin 6 (P6) revealed a very limited expression of P6 in the liver sections of normal animals, while there was a slight increase in the expression of P6 in hepatic cells of DEN-alone and solvent treatments (0-mg/kg group). No significant difference was observed in P6-positive stained cells in the livers of rats from different dose groups of GTP. The P6-positive cells elevated dramatically (p<0.05) only in the livers of LTP 40 mg/kg supplemented rats (Figure 6B, D).
Discussion
The antioxidant activity and hepatic cytoprotective effect of water-soluble GTP and lipid-soluble LTP was investigated in a two-stage model of rat hepatocarcinogenesis initiated with DEN and promoted by PB. DEN is thought to initiate hepatocarcinogenesis through production of reactive oxygen species (ROS). When the generation of ROS overwhelms antioxidant defenses, it has serious consequences including oxidative damage to key proteins, DNA mutations or adducts, and lipid peroxidation (29). Continuous administration of PB effects release of free radicals, together with DEN, strongly and rapidly predispose to hepatocarcinogenesis and drives liver tissue toward HCC. It has been observed that under our experimental conditions DEN initiation followed by PB promotion induced dramatic hepatic hepatocarcinogenesis in rats as evidenced by an increase in 8-OHdG levels as well as GST-P-positive foci, a sensitive and reliable marker of pre-neoplasia and neoplasia in hepatic tissues.
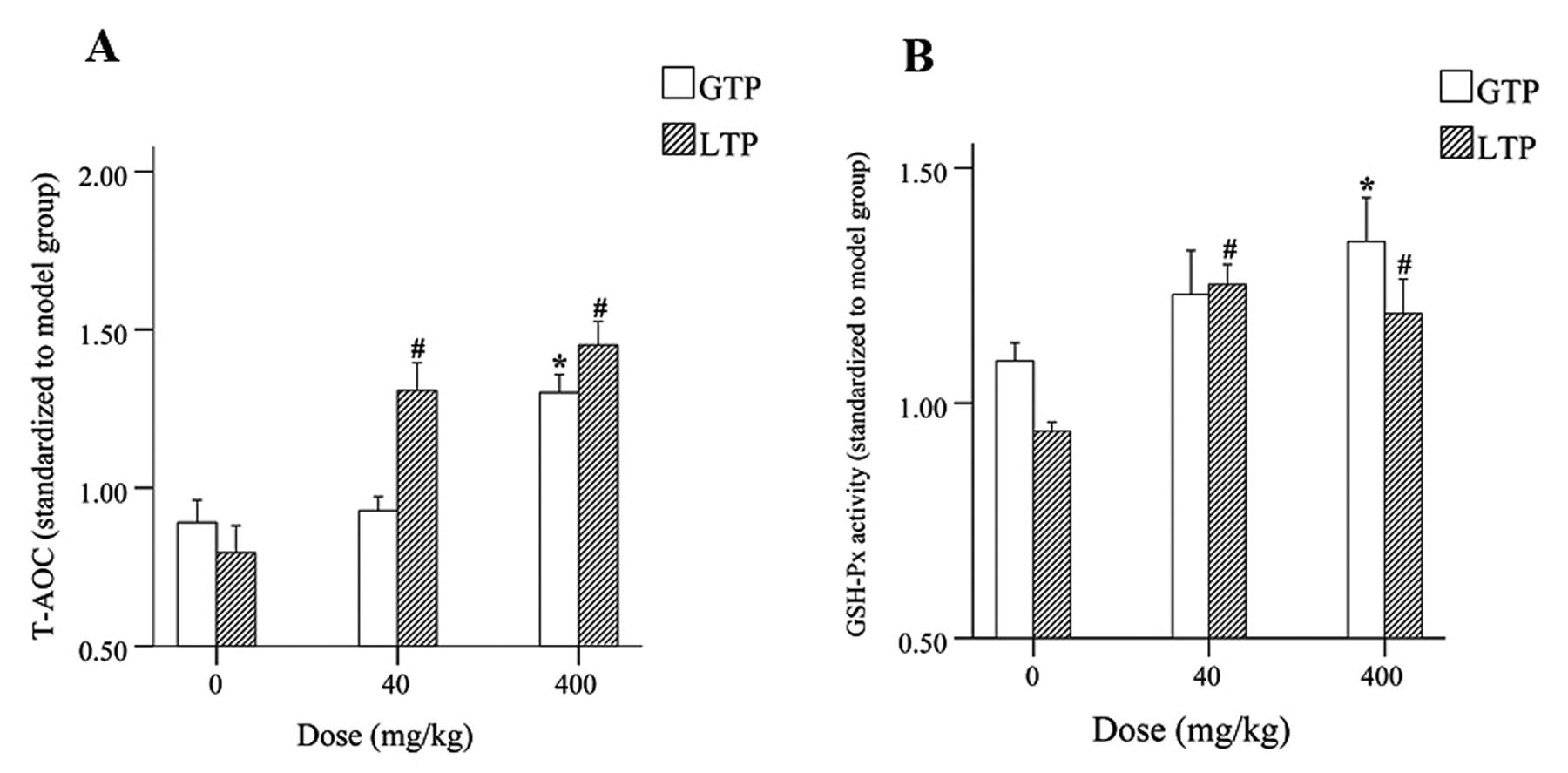
Antioxidant effects of GTP/LTP during DEN-induced hepatocarcinogenesis in rats. (A) Total antioxidant capacity in liver after exposure to GTP/LTP. (B) The activity of GSH-Px in liver after exposure to GTP/LTP. *p<0.05 compared to GTP 0 mg/kg group; #p<0.05 compared to LTP 0 mg/kg group. Data are shown as the mean±SE. Statistical significance was determined by ANOVA.
GTP possesses antioxidant and cancer chemopreventive activity in many different animal models of carcinogenesis (30-33), but the model for the antioxidant and anti-carcinogenic activities of LTP in chemically-induced tumorigenesis remains poorly-understood. In the current study, we present evidence, for the first time, that GTP/LTP mediated chemopreventive action through antioxidant activity against experimental hepatocarcinogenesis. Antioxidant enzymes such as catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GSH-Px), and glutathione S-transferase (GST) protect mammalian cells from oxidative stress, and consequently, reduce the propensity of tissues to develop malignancy. T-AOC was a representative of enzyme and non-enzyme antioxidant index in the liver. In our study, LTP exerts potent antioxidant activities as evidenced by a drastic increase in T-AOC levels and GSH-Px activity at both low and high doses. GTP demonstrated equivalent antioxidant activities only at higher dosages. The antioxidant activity may be prominent when high doses of GTP are ingested to counteract the effect of high levels of exogenous or endogenous ROS, however, rats exposed to high-dose GTP with DEN/PB experienced significant body weight loss. Similarly, there have been case reports showing that excessive amounts of GTP induce organ toxicity (34-36). Here, we provide important evidence that LTP possess better antioxidant activity and hepatic cytoprotective effect than GTP during DEN-induced early hepatocarcinogenesis.
To delineate the underlying mechanism of the antioxidant effect of GTP/LTP during DEN hepatocarcinogenesis, immunohistochemical (IHC) analysis was performed to monitor protein expression associated with oxidative stress. Surprisingly, little attention has been paid to the Nrf2 expression during DEN-induced hepatic damage. This information is important because a substantial proportion of cellular antioxidant enzymes, that neutralize oxygen-containing free radicals, are modulated by Nrf2 (37), which binds to ARE in DNA, thus increasing the expression of antioxidant enzymes. Nrf2 is well-regarded as a novel molecular target for chemoprevention (38). Our data show that the administration of GTP at a dose of 40 mg/kg resulted in larger numbers of Nrf2-positive stained cells compared to water solvent (0 mg/kg) treatment, whereas LTP at both 40 mg/kg and 400 mg/kg mediated a reduction of Nrf2 expression compared with oil solvent treatment. The former is easily explained because it has been previously reported that dietary antioxidants, e.g. pomegranate, resveratrol and black currant anthocyanins mediated chemoprevention of experimental hepatocarcinogenesis involves Nrf2-regulated antioxidant mechanisms (39-41). Nrf2-mediated antioxidant mechanisms may thus be important to the activity of GTP in alleviating oxidative stress during DEN hepatocarcinogenesis. On the other hand, based on our data, we speculate that activation of Nrf2 afforded by LTP may only be apparent in the presence of excessive oxidative stress. Once excessive oxidative stress is alleviated or disappears, the Nrf2 activation subsides and subsequently Nrf2 degradation initiates through associating with its cytosolic inhibitor Keap1 (42). It was similarly reported that the antioxidant effects of EGCG are only apparent in the presence of excessive oxidative stress as administering EGCG to old rats reduced oxidative stress, however, no effects were observed in young rats (43, 44). This LTP-induced decrease in the expression of Nrf2 has not been reported in the literature before. Thus, additional studies are required to determine whether there is a causal relationship between LTP administration and decreased Nrf2 expression.

Effects of GTP/LTP on antioxidant protein Nrf2 and P6 expression. (A) Representative immunohistochemistry microphotographs of Nrf2 in hepatic tissue. (B) Representative immunohistochemistry microphotographs of P6 in hepatic tissue. (C). Percentages of hepatic Nrf2-positive cells are shown in the graphs. Data are mean±S.E. (n=4, *p<0.05 compared to GTP 0 mg/kg group). (D) Percentages of hepatic P6 positive cells are shown in the graphs. Data are mean±S.E. (n=4, #p<0.05 compared to LTP 0 mg/kg group) (magnification: ×400).
P6 has been shown to be unique in its ability to reduce phospholipid hydroperoxides (45). It has also been shown that P6 is important in protecting tissue from various oxidative insults, owing to its glutathione peroxidase activity and its ability to reduce peroxidized membrane phospholipids (25). Previous studies have shown P6 to protect tissue from oxidative stress (45-47). Our aim was to investigate how the expression of P6 is affected by GTP/LTP during the early stages of DEN-induced hepatocarcinogenesis. In our study, no significant difference is observed in P6 protein levels in the livers of rats chronically-exposed to GTP. Unlike results in the GTP groups, significantly increased P6 expression was observed in IHC after exposure to LTP 40-mg/kg treatment. Up-regulation of P6 in disease models involving oxidative stress has not been widely reported. A previous study demonstrated that the expression of P6 was induced by hyperoxia during oxidative stress in both rat and mouse lungs (48). This is the first report of LTP-induced P6 expression behaving as an antioxidant and protecting the livers of experimental rats from ROS-induced hepatocarcinogenesis.
Conclusion
In summary, T-AOC together with GSH-Px activity serves as sensitive biochemical markers of anti-oxidative effects. Our data demonstrate the novel finding that LTP administration results in increased expression of P6 in the rat liver. Evidence from the experiments reported here support the conclusion that LTP treatment produces improved antioxidant activity and hepatic cytoprotective effects compared with GTP treatment during DEN-induced early hepatocarcinogenesis. Results from this study provide insights into the potential for alleviating oxidative injury through modulation of antioxidant protein as a potential strategy for liver cancer chemoprevention by LTP-derived products. Additional experiments are required to elucidate the underlying molecular antioxidant mechanisms mediated by LTP. This may provide promising opportunities in the prevention of DEN-induced hepatic damage and hepatocarcinogenesis.
Acknowledgements
This work was partly funded by grants from the National Natural Science Foundation of China (No. 81172659) and China Postdoctoral Science Foundation (No. 2013M531474), and was supported in part by Elizabeth & Anthony De Stefano Foundation.
- Received January 8, 2014.
- Revision received March 17, 2014.
- Accepted March 18, 2014.
- Copyright © 2014 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.