


Abstract
Background: The main aim of this work was to study the effects of single whole-body irradiation (WBI) on the lymphoproliferative response in radiation-sensitive BALB/c mice. Materials and Methods: Mice were irradiated (0-5 Gy) and euthanized immediately afterward; other animals were subjected to WBI (4 Gy) and were analyzed after periods of 0-180 days. Splenic cell number, lymphoproliferative response and lymphocyte subpopulations were studied. Results: This study shows that immediately after exposure, an inhibition of the basal mitogen lymphoproliferative response was produced; furthermore, B-cells appear to be more radiosensitive than T-cells. However, up to 90 day's post-irradiation, mice spleens clearly show low cell numbers, and subpopulations and T-cell mitogens did not return to normal, while the basal response and B-cell mitogens peaked on day 15 post-WBI. Conclusion: In our model, B-cells regenerate earlier than T-cells, while Th lymphocytes regenerate faster than Tc lymphocytes.
Ionizing radiation (IR) has a number of effects on the immune system, some of which can be serious if radiation is not controlled. For instance, survivors of high doses of radiation in war or nuclear accidents present numerous effects on their immune system long after the exposure (reviewed in 1, 2).
On the other hand, controlling the use of radiation can doubtlessly have beneficial medical effects despite the side-effects associated with it. Knowledge of these effects on the immune system in the short-, mid- and long-term is extremely important, not only for reasons of prevention, but also for administering medication before or indeed after IR exposure (3, 4).
At the molecular and cellular levels, IR may produce changes in nucleic acids, proteins, lipids and carbohydrates. Those affecting DNA and the enzyme systems required for recovery and apoptosis regulation, as well as the immune and hematopoietic systems which are particularly sensitive to IR, are of special importance (5, 6). For this reason, individuals exposed to whole-body irradiation (WBI) present a variety of symptoms affecting different organs or systems, and this situation occurs at low doses (7) with immune and hematopoietc systems.
The radiosensitivity of the immune system varies depending on the animal and breed being studied. This in the case of laboratory mice as the BALB/c strain is significantly more radiosensitive than others used in research, for example, C57BL/6J (8-10). Compared to other strains, this radio-sensitivity might relate to p53 pathway activity and apoptosis regulation (11), the expression of the Fas molecule and its receptor (12) and the lower production of IL-12 and IL-23 (13). This enhanced radiosensitivity of BALB/c is associated with these animals' increased susceptibility to develop spontaneous mammary tumors if compared to other mice strains (14).
As the BALB/c mouse strain is highly radiosensitive, it is striking that there is very little information available about the effect of ionizing radiation on the immune system in these animals, especially in the long term. These data will be of great importance for comparing the effects of ionizing radiation on the immune system because they depend significantly on the strain being used.
Lymphocytes are cells in a quiescent state (G0) that may be stimulated to enter a cellular cycle (G1) by antigens and mitogens. Therefore, we used mitogens specific to T-cells (concanavalin A, phytohemaglutinin A), B cells (bacterial lipopolysaccharide) and mixed (pokeweed mitogen), which consequently give rise to activation, proliferation and cell differentiation (15, 16). However, lymphocytes constitute a heterogeneous mixture of subpopulations: CD3+ T lymphocytes (further separated into CD4+ T helper (Th) and CD8+ T cytotoxic (Tc)), B lymphocytes and natural killer (NK) cells. Sensitivity to IR changes in accordance with the B>T> NK subpopulations and the Tc>Th cells (17-19), and apparently relates to the differences in DNA repair mechanisms (20, 21).
Nonetheless, results for WBI are scarce, especially concerning ‘long-term effects’ and BALB/c strain mice. Thus, exposure of C57BL/6J mice to different doses of radiation in vivo brings about various immunological anomalies in vitro. Exposure to 2 Gy leads to even higher concentrations of lymphocytes and erythrocytes after 110 days (18), 3 Gy exposure results in not only an anomalous secondary immune response with LPS, but differences in lymphocyte populations (22). Increased numbers of T CD4+ lymphocytes have also been observed a few days after exposure (23), as have reductions in the number of helper Th1 cells with the consequent altered number of Th1/Th2 (23), diminished IL-12 production by the T lymphocytes (24), and differences in the NK response relating to the Th1/Th2 balance (25, 26).
One accepted method to evaluate immunocompetence is the study of lymphocyte proliferation in response to mitogen stimulation. The use of distinct mitogens specifically for the distinct lymphocyte populations enables us to obtain an immunocompetence value of an organism, and for any alterations taking place such as the types of cells affected and the possible mechanisms involved. However, this method is only rarely used and tends to be applied specifically. Some authors have described inhibitions of both spontaneous cell division and the cell division induced by mitogens. The degree of inhibition varies according to the mitogen used and the target cell (27, 28). What is more, there are very few data available on long-term changes.
For these reasons, an immunocompetence study after in vivo exposure for a radiosensitive strain such as BALB/c, measured in terms of the lymphocyte proliferation induced by a wide range of mitogens, coupled with prolonged long-term observation, is more than desirable. In this work, we studied the immediate, short-, mid- and long-term effects of in vivo exposure on the lymphoproliferative response induced by mitogens in radiosensitive BALB/c mice, and we discuss the possible mechanisms of the effects observed.
Materials and Methods
Animals. Male and female BALB/c mice were used, which were free of pathogens, weighed 22-25 grams and were 8 to 12 weeks old. They were bred in the research center and kept in plastic boxes (4 mice per box) at a constant temperature (24°C±1°C) and relative humidity (75%±5%). They were fed on rodent food (Purina Mouse Chow, Valencia, Spain) and were provided water ad libitum.
Whole-body gamma-irradiation in vivo procedure. Animals were exposed to gamma radiation, electromagnetic radiation by photons with energy propagation. In vivo irradiation took place in a Sagitarius (25 MV) linear accelerator at the Oncology Service of La Fe Hospital, Valencia. The dose rate was 2.4 Gy/min, with a dose error under 2%. Mice were placed individually in ventilated methacrylate boxes whose structure held the animals in a supine position.
After irradiation, mice were either killed immediately to study the immediate effects of radiation or transferred back to the laboratory to study the short-, mid- and long-term effects. All the animals were rapidly and humanely euthanized with 100% CO2 in accordance with standard international animal care protocols (29).
To study the immediate effects of WBI on the lympho-proliferative response, five groups of mice were chosen (12-14 per group). They were administered a single dose of WBI (0, 1, 3, 4, or 5 Gy) and killed immediately afterward.
To study the short-, mid- and long-term effects of the lympho-proliferative response, six groups of mice were given a single dose of WBI (4 Gy), were killed and the proliferation test was carried out after periods of 0, 7, 15, 30, 90, and 180 days.
We considered the effects observed at 0, 7, 15, and 30 days short-term effects, those noted after 90 days were taken as mid-term effects, and long-term effects were considered to take place after 180 days.
Preparation of spleen cell suspensions. The spleens of both the irridated mice and the control group were removed under sterile conditions, and cell cultivations were prepared using sterile pressure sieves. Cells were washed twice in saline solution and then resuspended in RPMI-1640 (Gibco, Paisley, Scotland) containing 5% of fetal calf serum (Gibco), glutamine 2 mM (Flow Laboratories, Irvine, Scotland) and 100 μg/ml cefotaxime (Hoechst, Barcelona, Spain) at a density of 2×106 cells/ml.
Proliferation cell assay. The spleen cells of the irradiated animals following the two aforementioned procedures, while those of the control group were placed in 96-well plates with 4×105 cells/well where they were cultivated without mitogens or stimulated with different doses of concanavalin A (ConA; Sigma-Aldrich, Spain), phytohaemaglutinin A (PHA; Sigma-Aldrich), lipopolysaccharide from Ercherichia coli (LPS; Difco, Spain) and pokeweed (PWM; Gibco).
Cultures were prepared in duplicate and incubated at 37°C in a humid atmosphere with 5% CO2 for 48 hours. Next, 18.5 kBq of tritiaded thymidine (185 GBq or 5 Ci/mmol, Amersham Bioscience, Spain) were added to each well; thus, each culture received 0.5 mCi. Cells were collected 18 hours later by suction through fiberglass filters and the radioactivity in the cells was measured with a liquid scintillation counter.
It is well-known that changes occur in the lymphoproliferative response throughout the short life of BALB/c mice, thus rendering the use of control groups necessary. The first control group (C1) was used as a reference to study both the immediate effects (0 days) and short-term effects (7, 15 and 30 days post-irradiation) because there is no difference in the lympho-proliferative response over a 1-month period. In each stage, 4 mice were euthanized until a single control group was formed and made up of 16 mice, whose average proliferation value was taken as the reference point. For the study of the mid-term effects (90 days), a second control group was used (C2) which comprised 8 mice of the same age that were euthanized at the same time as those irradiated for this second stage. A third control group (C3), also formed by 8 mice, was used in the same way and as a reference point for the study into the long-term effects (180 days).
Spontaneous and mitogen-induced immunoproliferative responses in non-irradiated (Gy=0) and irradiated mice (Gy=1–5). Arithmetic mean±standard deviation of counts per minute are shown. Significance (unpaired Student's t-test): a=p<0.05, b=p<0.01, c=p<0.001, d=p<0.0001. N=number of animals.
Study of cell viability. Other than the previously-described cultivations, a study of cellular viability was conducted with the irradiated and non-irradiated animals using the cytometric acridine orange method (30).
Study of lymphocytes subpopulations. The study of lymphocyte subpopulations was carried out using specific antibodies which are marked and read by cytofluorimetry (Beckman Coulter, Spain). Basically, B lymphocytes were identified by a standard direct immunofluorescence test. The cell pellet was incubated with fluorescein-conjugated goat anti-mouse Ig for 30 min at 4°C and washed; cells were then fixed in 0.5 ml ethanol/0.5 ml saline solution for the subsequent flow cytometry analysis using an EPICS-PROFIlE flow cytometer. Thus, the T lymphocyte set and subsets were identified by the indirect immunofluorescence test. Cell pellets were incubated with anti-Thy1, anti-Lyt2 (reacting with Tc cells) or anti-L3T4 (reacting with Th cells) monoclonal antibodies (rat IgG2b anti-mouse; Sera-Lab, Spain) for 30 min at 4°C and were then washed. Cells were stained by incubation with a second fluorescent monoclonal mouse anti-rat IgG2b-fluorescein isothiocyanate (FITC)-conjugated antibody (Sera-Lab) for 30 min at 4°C. After washing, cells were resuspended and fixed in 0.5 ml ethanol/0.5 ml saline solution for the subsequent flow cytometry analysis.
The results of the lymphocyte subpopulations were compared in the euthanized mice at the same times as the control group in each case.
Statistical analysis. A comparison of the proliferation tests was made of the results obtained between the irradiated and non-irradiated animals with Student's non-paired t-test. In both the cellular viability and the lymphocyte subpopulation studies, comparisons with the control group were made using a Mann-Whitney nonparametric test.
Results
Immediate effects of WBI on the lymphoproliferative response. As shown in Table I, IR in vivo inhibits both spontaneous DNA synthesis and that induced by mitogens in mouse spleens. This inhibition depends on the WBI dose administered to the mouse.
We can see differences in the degree of variation and the level of significance which depends on mitogen-induced proliferation. Thus those spleen cells stimulated with B-cell mitogens (LPS or PWM) were significantly inhibited with the 1 or 3 Gy WBI doses, or with higher ones. Yet those cells treated with ConA or PHA were only significantly inhibited when the radiation doses administered either equalled or exceeded 4 Gy.
By considering these results, a decision was made to use 4 Gy doses for the rest of the experiments as it was the lowest WBI dose which significantly inhibited the cellular proliferation induced by all the mitogens under study.
The immediate, short-, mid- and long-term effects of WBI on the lymphoproliferative response.
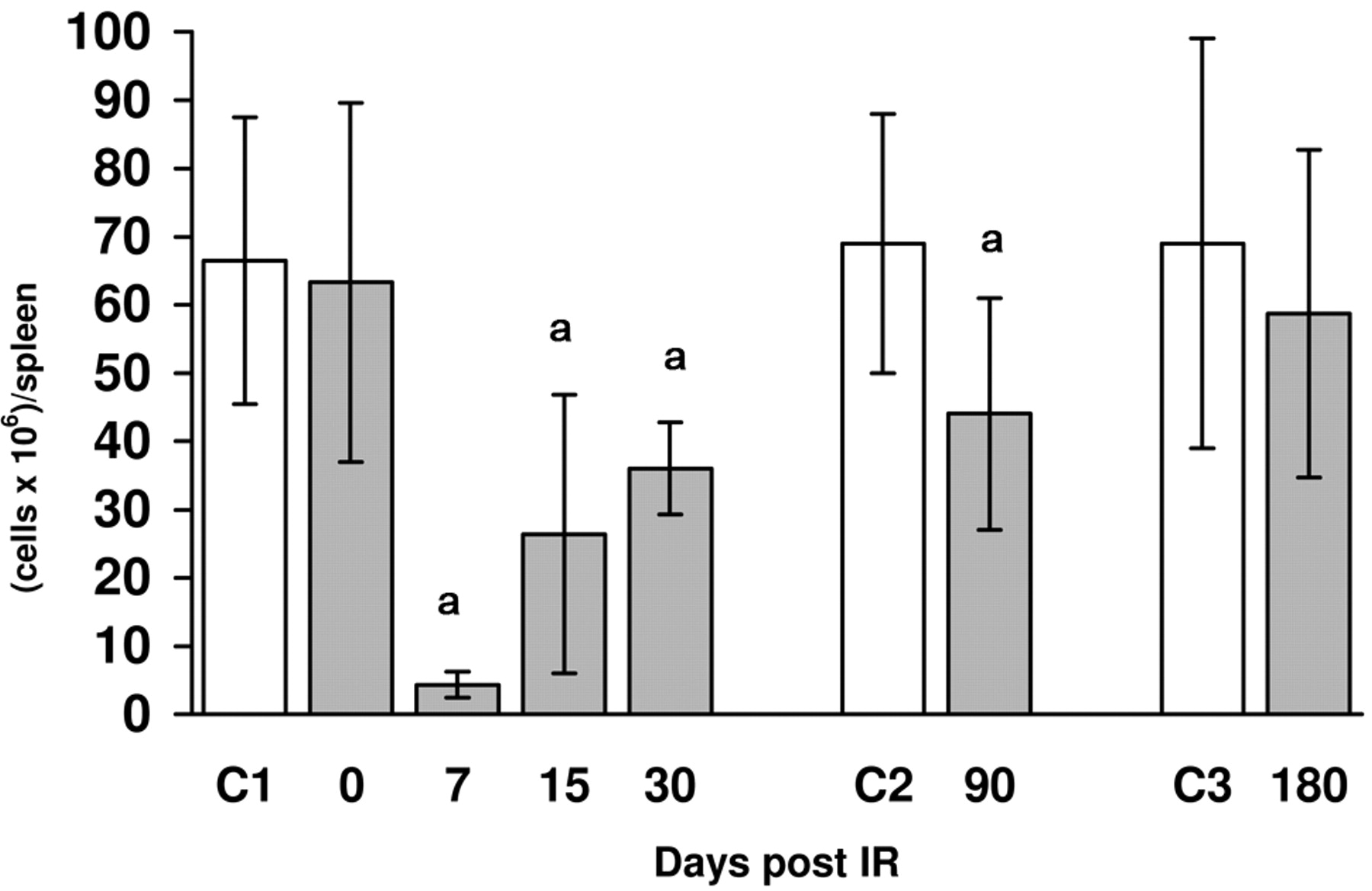
A) Number of cells: Figure 1 shows the number of spleen cells taken from mouse spleens after irradiation at 4 Gy. A sharp drop in spleen cell numbers was noted, which reached its lowest point on day 7 post-exposure. Spleen cells increased gradually until they reached almost normal levels on day 180. B) Lymphocyte subpopulations: The study of lymphocyte subpopulations is presented in Table II. Immediately after irradiation, there were no significant variations in the percentages of the B, T, Th, or Tc lymphocytes. On the other hand, the percentage of B-cells dropped significantly. The percentage of T-cells remained constant even though a marked increase in the quotient Th/Tc was seen, which was caused by a significant drop in the Tc cells accompanied by an increase in Th cells. On day 15 post-irradiation, a percentile depletion of T and the Th and Tc lymphocytes was seen, although the Th/Tc ratio was higher than that of the control group. Moreover, a significant increase in B lymphocytes was also observed. However on day 30 post-irradiation, the percentage of total T-cells significantly lowered without affecting the proportion of Th and Tc cells, while B-cells were normal. The parameters were normal after 90 or 180 days, although the percentage of B-cells slightly increased. C) Activation of lymphocytes: Immediately after irradiation, spontaneous DNA synthesis (Figure 2) was also inhibited significantly, but increased later. At 15 days, in irradiated animals DNA synthesis was higher than that of the control group.
Effects of a single dose of ionizing radiation in vivo exposure (4 Gy) on lymphocyte percentages and Th/Tc ratio in respect to post irradiation time. Arithmetic mean±standard deviation is shown for each type of lymphocyte. N=number of animals. Significance (Mann Whitney test), a=p<0.05.
As Figures 3 and 4 depict, a single WBI dose of 4 Gy significantly inhibited the DNA synthesis induced by ConA or PHA immediately upon irradiation and for the following days. Proliferation ability gradually recovered at 30 days post-irradiation to register a similar value to that of the control group. Moreover at 90 and 180 days, the ConA-induced DNA synthesis was significantly higher in the irradiated animals if compared immediately after irradiation, but increased later.

Effects of a single dose of WBI (4 Gy) on the splenic cell number isolated 0, 7, 15, 30, 90 and 180 days post-irradiation. Arithmetic mean±standard deviation is shown. Annimal numbers euthanized were 95, 39 and 15 for the control groups C1, C2 and C3 respectively, and 30, 25, 42, 25, 27 and 12 for the irradiated groups at 0,7, 15, 30, 90 and 180 days. Significance (Mann Whitney test): a=p<0.0001.

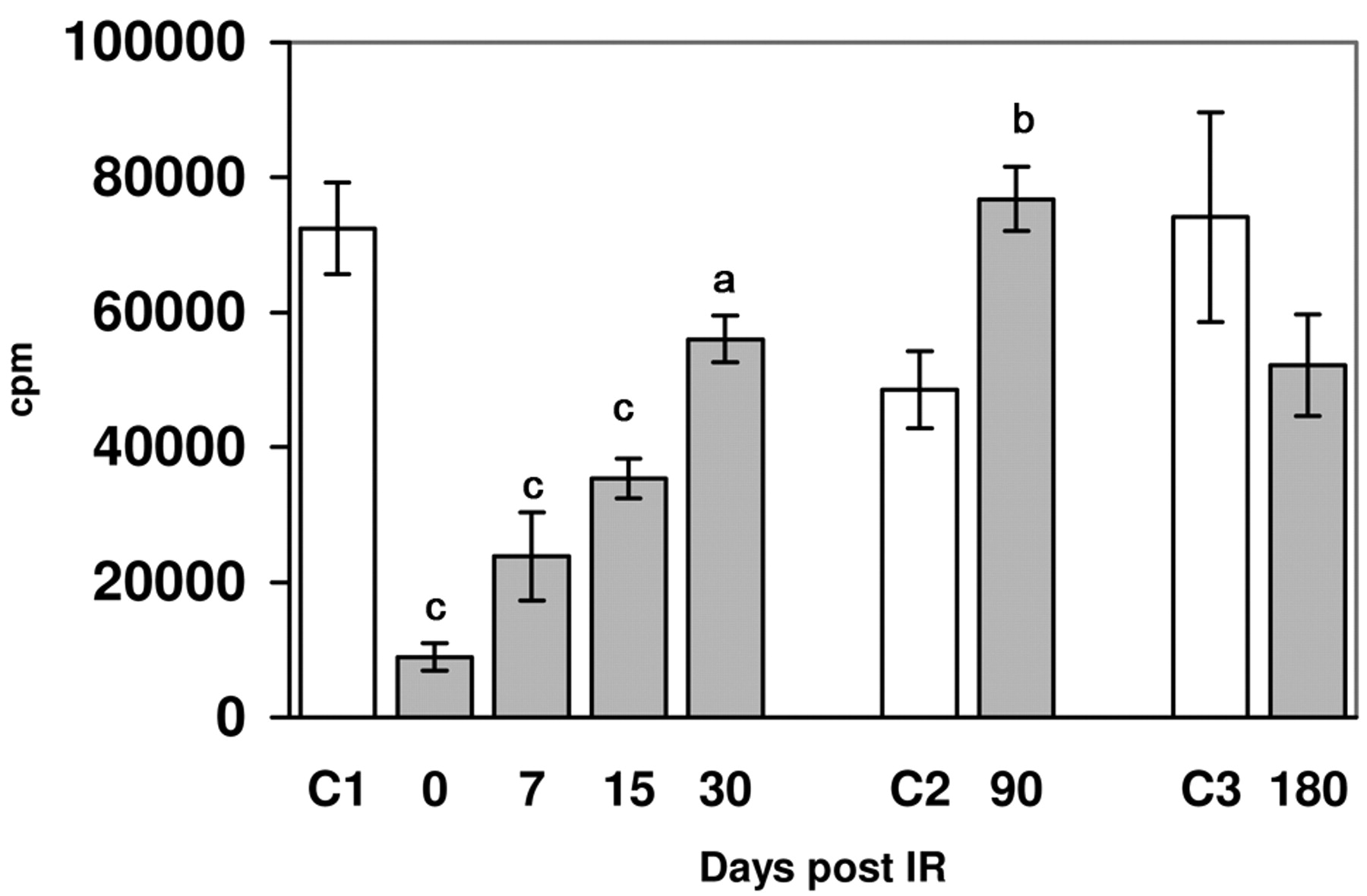
Basal lymphoproliferative response in control groups (C) and irradiated mice in respect to the time post irradiation. Arithmetic mean±standard deviation is shown. Annimal numbers euthanized were 30, 15 and 12 for the control groups C1, C2 and C3 respectively, and 26, 25, 29, 25, 26 and 12 for the irradiated groups at 0, 7, 15, 30, 90 and 180 days. Significance (unpaired Student's t-test): a=p<0.001, b=p<0.0001.
Spontaneous DNA synthesis (Figure 2) and the proliferation induced by LPS or PWM (Figures 5 and 6) followed a similar response pattern after radiation inhibition, and both reached their highest value on day 15 post-exposure. That is, those cells that repopulate the spleen showed a peak on day 15 post-irradiation, irrespectively of them being stimulated or not.
Thus a significant inhibition took place immediately after irradiation, but returned to normal later to reach significantly higher values than those recorded in the control group on day 15, and finally lowered to normal values thereafter.
There were no appreciable differences in cellular viability between the irradiated and non-irradiated animals as both groups obtained values of around 90% in all cases (data not shown).

Lymphoproliferative response induced by 1 μg/ml of concanavalin A in control groups (c) and irradiated mice in respect to the time post irradiation (similar results were obtained with 3 μg/ml of concanavalin A). Arithmetic mean±standard deviation is shown. Annimal numbers euthanized were 30, 15 and 12 for the control groups C1, C2 and C3 respectively, and 26, 25, 29, 25, 26 and 12 for the irradiated groups at 0, 7, 15, 30, 90 and 180 days. Significance (unpaired Student's t-test): a=p<0.05, b=p<0.001, c=p<0.0001.
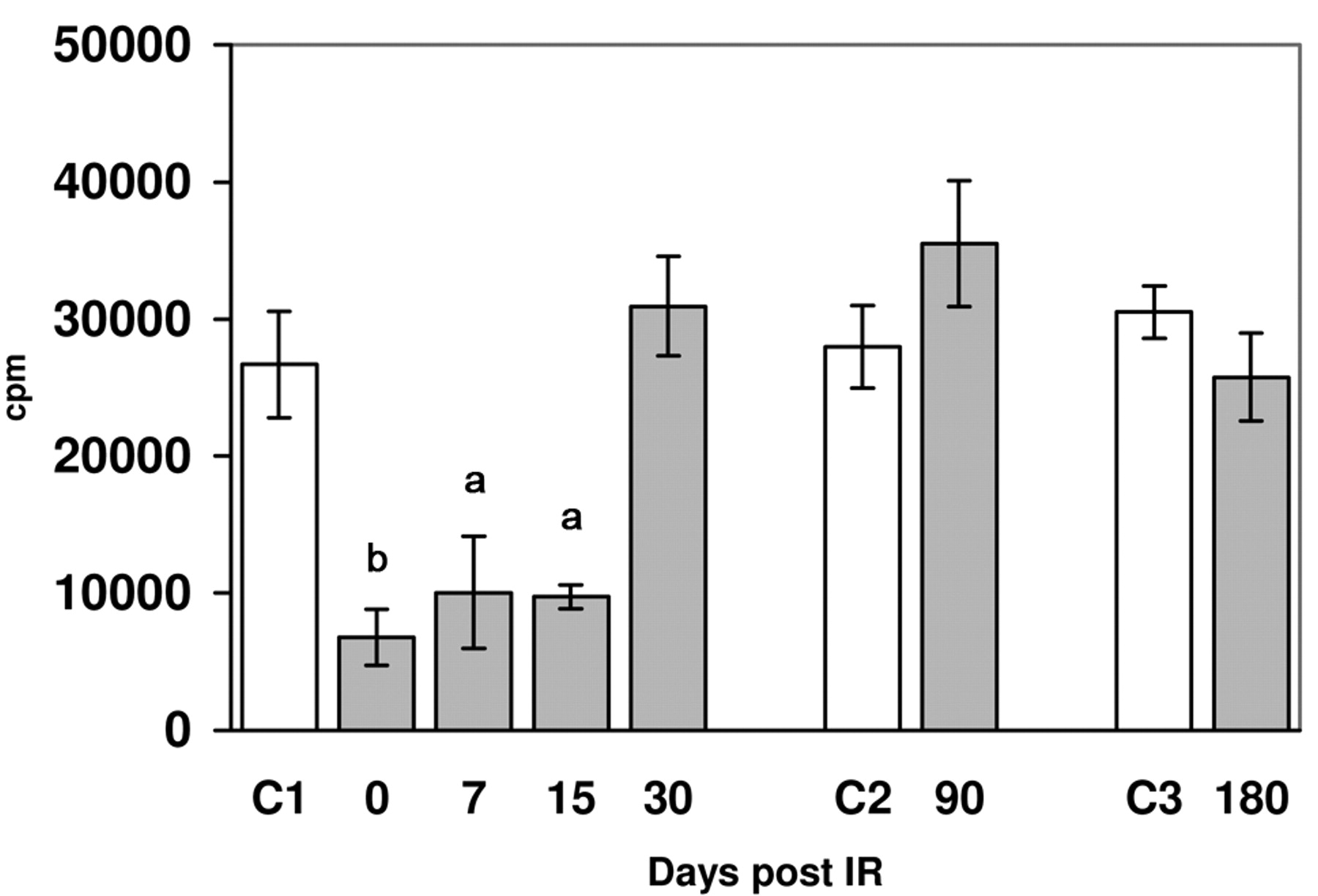
Lymphoproliferative response induced by 10 μg/ml of phytohaemaglutinin A in control groups (c) and irradiated mice in respect to the time post irradiation (similar results were obtaines with 5 μg/ml of phytohaemaglutinin A). Arithmetic mean±standard deviation is shown. Annimal numbers euthanized were 30, 15 and 12 for the control groups C1, C2 and C3 respectively, and 26, 25, 29, 25, 26 and 12 for the irradiated groups at 0, 7, 15, 30, 90 and 180 days. Significance (unpaired Student's t-test): a=p<0.001, b=p<0.0001.
Discussion
The mammal cell response to IR is determined genetically and mouse strains can be identified by their radiosensitivity. The reason for choosing BALB/c mice as opposed to other strains (for example C57BL/6J) is because they have proven to be exceptionally sensitive to IR (8-10). In addition, the use of the immune system allows us to observe molecular and cellular alterations, if they indeed take place.

Lymphoproliferative response induced by 40 μg/ml of lipopolysaccharide in control groups and irradiated mice in respect to the time post irradiation (similar resuls were obtained with 80 μg/ml). Arithmetic mean±standard deviation is shown. Annimal numbers euthanized were 30, 15 and 12 for the control groups C1, C2 and C3 respectively, and 26, 25, 29, 25, 26 and 12 for the irradiated groups at 0, 7, 15, 30, 90 and 180 days. Significance (unpaired Student's t-test): a=p<0.001, b=p<0.0001.

Lymphoproliferative response induced by 1% of pokeweed in control groups and irradiated mice in respect to the time post irradiation. Arithmetic mean±standard deviation is shown (similar results were obtained with 2%). Annimal numbers euthanized were 30, 15 and 12 for the control groups C1, C2 and C3 respectively, and 26, 25, 29, 25, 26 and 12 for the irradiated groups at 0, 7, 15, 30, 90 and 180 days. Significance (unpaired Student's t-test): a=p<0.05, b=p<0.001.
Previous studies have demonstrated that such high doses of WBI (4 Gy) cause major atrophies in the spleen with a very slow recovery (31, 18, 10). In our model, recovery of cell numbers only takes place after 180 days; therefore, an increase in cell proliferation is required, which we can see in Figure 2, particularly the data corresponding to days 7-30 post-irradiation. Our results are similar to those of others (31, 18, 10) who state that the effects of WBI on the murine hematopoietic system can last for long periods and that radiation may also affect mouse strains in accordance with their genomic instability. Thus, BALB/c strain mice are more sensitive to radiation than are the C57BL/6 strain mice with which most authors have worked in both the short and long term (10). Aberrations were noted up to almost 180 days after exposure to radiation; in humans, this period can be 24 years in some cases, and survivors of the atomic bomb were harmed proportionally to the irradiation levels recorded after a 60-year period (1).
It is well known that treating mice and humans with IR induces a serious depletion in the number of lymphocytes, as well as alterations in the percentages of the different subpopulations of these cells: in general, B-cell subpopulations are more radiosensitive than T-cells, and both are more radiosensitive than NK cells (17-19). However, this is not the case of the NK function; for example, C57BL/6 mice received a dose of gamma-WBI (4 Gy) and presented a deficit in the secretion of interferon-gamma over long periods (26). What is more, the immunology of the survivors of the atomic bomb reveals significant changes, that is, a decrease in the Th cell populations and an increase in the blood inflammatory cytokine levels (reviewed in 1). Therefore in our model, BALB/c mice are much more susceptible to infection than C57BL/6 mice, and they generally display deficiencies in Th1 cytokines and lymphocytes (13).
Our results show that after WBI (4 Gy), significant changes take place in the distinct spleen lymphocytes subclasses, thus, the percentage of T-cells does not change (Table II, control=44±9 vs. 7 days post-irradiation=53±8) even if the number of spleen cells (Figure 1). On day 7 post-irradiation, a relative reduction in B-cells is seen, while the number of T-cells remains unchanged. However, a spectacular increase in the Th/Tc ratio is observed, with a reduction in Tc cells and a percentile increase of Th cells. Once more, these results suggest that B-cells are more radio-sensitive than T-cells and that Tc cells are more radio-sensitive than Th cells. At day 15 post-WBI (when the recovery process of spleen cells has already begun), a significant increase of B-cells is noted with a reduction in T-cells, while the Th/Tc ratio is higher than that in the control group. On the other hand, the results on day 30 post-WBI are practically normal as regards the percentage of cells, although a slight reduction of T-cells may be seen. Lastly, Table II shows how B-cells deviate markedly from the control group on days 90 and 180 post-WBI. These results are similar to those reported by other authors (18, 31, 32) who used the mouse strain C57BL/6 which is subject to WBI (0.5 and 3 Gy) with euthanasia at 4 or 110 days. Other authors used C57BL/6 mice and demonstrated how treatment with 0.1 and 0.7 Gy with blood samples taken on days 1, 4, and 7 post-WBI induces a significant increase in CD4+ T lymphocytes and that B-cells are more sensitive to radiation (23). On the other hand, an increase in the Th/Tc ratio indicates an inflammatory response (33, 34). In addition, ionizing radiation is known to reduce Th1 and, consequently, the Th1/Th2 imbalance, and the C57BL/6 mice received a dose of gamma-WBI of 4 Gy and were immediately killed. These authors' findings reveal that the NK1.1+ T-cells, which survive in an early post-irradiation stage, play an important role in the Th1/Th2 balance (25). Conversely, the mice receiving a gamma-WBI on day 50 post-irradiation show low levels of Th1, consequently leading to a Th1/Th2 imbalance. These results indicate that the NK cell is an important factor in the lessened Th1-like response in irradiated mice (26). Therefore, we may state that our mice lymphocytes display a unique behavior, the Th/Tc ratio is remarkable on days 7-15 post-irradiation, and the Th1/Th2 ratio is above average with very low Th1 lymphocytes at the time of irradiation. For these reasons, we may state that the percentile normalization of the distinct lymphocyte subclasses precedes the recovery of the number of cells per spleen in temporal terms with an earlier spleen repopulation than both the B- and T-cells.
Proliferation: immediate effects. Table I shows how WBI causes an immediate inhibition of both the spontaneous proliferative responses and that induced by specific mitogens (ConA, PHA, PWM and LPS). The resulting inhibition is dependent on the IR dose. These results are similar to those obtained by other authors (35, 36). Moreover, we observed that the differences in the degree of inhibition seem to depend on the type of mitogen proliferation inducer. Thus the responses induced by LPS or PWM are inhibited by lower IR doses than that depending on ConA or PHA (Table 1). These results obtained after in vivo irradiation, together with the results of the in vitro irradiation studies, show that B cells are more radiation-sensitive than T-cells. This finding is supported by the results reported by other authors (17, 21, 28). However, we have observed that the effects of different IR doses on lymphocyte proliferation do not depend on an immediate decrease in viability (28).
Proliferation: short-, mid- and long-term effects. The results obtained in this work demonstrate a marked increase in spontaneous DNA synthesis 15 days after irradiation, and the control values multiplied 3-fold, an increase which was maintained at 30, 90 and 180 days, although it was not significant. There are significant increases in spontaneous proliferation under experimental conditions. Generally, these increases can be explained as the result of the repopulation of the spleen, and by the fact that this organ in adult mice and humans possesses hematopoietic capacity (37, 38, 1).When viewing the figures of non-stimulated cells (LPS and PWM), it is interesting to note that they may overlap, however, the same cannot be said for non-stimulated cells (ConA and PHA). Therefore on day 15 post-irradiation, cells divide, particularly B-cells (LPS), as do the PWM stimulated cells, but to a lesser extent; this is certainly one of the most innovative findings of this work.
Lastly the behavior of the cells stimulated by B-cell mitogens (LPS and PWM) is the same as that of the non-stimulated and irradiated cells. They were normal on day 7 post-IR. However on day 15 post-WBI, the increase of DNA synthesis was double (LPS). Lastly from day 30 post-WBI, the results are the same for both irradiated mice and the control group. As the proliferative response of B dependents was normal on day 7, this coincides with a significant decrease in the percentage of B lymphocytes, which makes us think about a possible hyperfunctionality of these cells at the time the analysis was made. Hyperfunctionality could be caused by either a decreased percentage of T-cells with a suppressing function and or an increased percentage of collaborating T-cells. This situation becomes clearer with the responses induced by PWM as this mitogen requires the collaboration of Th cells to induce the proliferation of B-cells (39, 40). This explains why on day 15 post-irradiation, a hyperresponse to PWM paralleling that observed for LPS was not seen; instead a decreased percentage of T-cells inhibited a parallel increase in the response induced by PWM.
T-Cell mitogens (ConA and PHA) behave very differently in DNA spontaneous synthesis and in both LPS and PWM. Thus, the response induced by ConA and PHA gradually recovers with time and becomes practically normal on day 30 post-irradiation, but without a peak which is so characteristic on day 15 post-WBI.
After 7 days of treatment with IR, and coinciding with an enhanced proliferative response of T-dependents, a sharp drop in the percentage of Tc lymphocytes and an increase in the Th/Tc ratio take place. These alterations are still noticeable, be it slightly, after 15 days (25). By taking into account previous results in mouse spleen cells that demonstrate a marked, significant interdependence between the intensity of the proliferative responses induced by mitogens and the percentage value of Tc cells (27, 41, 42), it seems that the depression in responses as opposed to ConA and PHA observed in the two weeks after irradiation is due to a drop in the percentage of Tc lymphocytes. In fact, the percentage nomalization of Tc cells and the Th/Tc ratio observed on day 30 coincide with a parallel normalization of the T-cell response capability when confronted with mitogens ConA and PHA (27).
Using a mouse model which is highly sensitive to radiation, our results suggest the following: a) irradiated mouse spleens are much smaller than those of the control group and continue to be so up to 90 days; b) cell subpopulations are highly modified by a single dose of WBI: on day 7, B lymphocytes are almost nonexistent and the Th/Tc ratio has a value of 15, yet on days 15 and 30 days, a complete change takes place and B-cells are the majority, the Th/Tc ratio is 4.67 and 3, respectively and it does not normalize until day 90; c) the effect of spontaneous lymphocyte proliferation is very revealing: on day 15 post-WBI, it was almost triple that of the control group; d) as for mitogens, the proliferation that takes place for B-cells follows a characteristic pattern, reaching peaks on day 15 post-irradiation, which differs from T lymphocytes since they reached their peak on day 90 post-WBI.
Our results, like those of other authors (1, 43, 44), suggest that our model based on BALB/c mice that are much more sensitive to gamma-WBI, in the spleen, other than those of hematopoietic stem cells, B-cells regenerate before T-cells do, while Th lymphocytes also regenerate more quickly than Tc lymphocytes. Our results testify to the immune system's ability to eventually recover normality after stimulation has finalized.
Acknowledgements
The Authors thank Dr. A González-Molina and the entire staff of Experimental Immunology Unit (Hospital La Fe, Valencia, Spain). This work was supported in part by a grant P.I.E. 04428.
- Received November 25, 2009.
- Revision received June 2, 2010.
- Accepted June 8, 2010.
- Copyright © 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}