


Abstract
Background: Recent data have demonstrated that mesenchymal stem cells (MSCs) have potent immune regulation capacity in vitro, enhancing their therapeutic appeal for their utilisation in the management of acute graft-versus-host disease (aGvHD). However, their immunoregulatory activity in vivo is largely unknown. Materials and Methods: Using murine compact bone-derived MSCs in an aGvHD model, the phenotypic status of splenocytes of aGvHD and aGvHD+MSC groups of mice were examined by flow cytometry. Results: MSC infusion decreased the expression of MHC-II and CD69 molecules on splenic CD11b+ cells of aGvHD mice, which resulted in decreased maturation of antigen-presenting cells. Moreover, the down-regulated ratio of CD3+CD69+ to CD3+ cells, leading to restrained early activation and effector T-cell formation, resulted in the enhancement of the absolute and relative number of splenic CD3+ cells by MSCs co-transfer. Conclusion: This study demonstrated that MSCs can inhibit the three developmental stages of aGvHD.
- Acute graft-versus-host disease
- antigen presenting cells
- immunosuppressive
- in vivo
- mesenchymal stem cells
- T lymphocyte
Mesenchymal stem cells (MSCs) were first recognised in the late 1960s by Friedenstein et al., who identified an adherent, non-phagocytic, fibroblast-like population that could regenerate rudiments of normal bone in vitro and in vivo (1-4). MSCs have been isolated from most non-human species such as mice (5), pigs (6), canines (7), rabbits (8), felines (9) and rats (10). Apart from bone marrow, MSCs have also been found in other postnatal tissues, such as placenta, adipose muscle, scalp tissue, brain, spleen, liver, kidney, lung, thymus, pancreas, as well as in various foetal tissues (11).
The immune phenotype of MSCs (widely described as MHC-I+, MHC-II−, CD40−, CD80−, and CD86−) is regarded as non-immunogenic. Furthermore, many reports have also described that MSCs have potent immunosuppressive properties. MSCs can modulate T lymphocyte proliferation induced by allo-antigens in mix lymphocyte reaction (MLR) in a dose- and time-dependent way (12). The suppressive activity of MSCs was greatest when added at the beginning of the MLR (13). Moreover, it has been shown that MSCs have immunomodulatory properties impairing maturation and functionality of dendritic cells (14) and B lymphocytes (15). There are also some reports that administration of MSCs can significantly prolong the mean survival time (MST) of MHC-mismatched skin grafts in baboon (16) and murine models (5).
Allogeneic haemopoietic stem cell transplantation (HSCT) is the most efficient treatment for many haematological malignancies and primary immunodeficiencies. However, the transplants also contain mature donor T-cells, which, when reacting with recipient histoincompatible antigens, can induce acute graft-versus-host disease (aGvHD), a life-threatening complication of allogeneic HSCT. aGvHD can be conceptualised as a three-stage process consisting of: (i) the initiation of tissue damage, (ii) the activation and proliferation of donor T-cells and (iii) the effector phase, resulting in tissue damage (17).
Owing to their potent immunoregulatory capacity, MSCs have recently received centre stage attention in the clinical setting to treat aGvHD. A number of successful clinical trials have shown the promising prospect of the use of MSCs in the treatment of aGvHD (18-19). Due to the use of different methods and different species to generate murine MSCs, conflicting results of MSCs in the murine aGvHD model have been produced (20, 21), limiting the investigations of the exact mechanisms by which transplanted MSCs act in vivo.
An understanding of the mechanisms by which MSCs inhibit aGvHD is critical for optimising this therapeutic approach. Using murine compact bone-derived MSCs in a murine aGvHD model, this study addressed this question by comparing phenotypic changes of splenocytes that occur in aGvHD mice with those of aGvHD mice receiving MSC transfusion.
Materials and Methods
Mice. Normal inbred C57BL/6 (H2b) and BALB/c (H2d) female mice were obtained from the Laboratory Animal Center, Academy of Military Medical Sciences (Beijing, China), and were housed in conventional cages. All experiments in this study were performed in accordance with the Academy of Military Medical Sciences Guide for Laboratory Animals.
Culture of murine bone-derived MSCs. MSCs derived from murine compact bone were isolated and culture-expanded as described previously (5). Briefly, long bones from female C57BL/6 mice aged 2-3 weeks were digested by collagenase II (Sigma-Aldrich, St. Louis, MO, USA) and the remaining bone debris was cultured in alpha-minimal essential medium (α-MEM; Gibco, Grand Island, NY, USA) containing 10% foetal bovine serum (FCS, Stem Cell Technologies, Vancouver, BC, Canada) in a humidified atmosphere of 5% CO2. The adherent cells at passage 3-5 were used in the experiments described below.
Murine aGvHD model (22). Nucleated splenocytes from female C57BL/6 mice were prepared after lysing RBCs with 0.84% NH4Cl and an inoculum of 7×107 viable cells without (aGvHD group) or together with 1×106 MSCs (aGvHD+MSC group) in a total volume of 0.25 ml PBS was intravenously injected into sublethally irradiated (5 Gy) BALB/c mice (n=40). Specimens from at least three recipient mice per group were collected for experiments except where otherwise described.
Flow cytometry (FCM) analysis. For detecting the immuophenotypic status of MSCs, expanded MSCs at passage 3 were collected and reacted with FITC- or PE-conjugated monoclonal antibodies against mouse MHC-I, MHC-II(I-Ab) (eBioscience, San Diego, CA, USA) for 30 minutes in the dark. The percentage of MSCs in each cell cycle phase was assessed by propidium iodide (PI) incorporation. To detect the phenotypic status of splenocytes of the aGvHD and aGvHD+MSC groups of mice, 1×106 nucleated cells per spleen at various observation times after transplantation were reacted with FITC- or PE-conjugated monoclonal antibodies against mouse CD11b, CD69, MHC-II, CD3 (eBioscience) for 30 minutes in the dark. The stained cells were washed twice in cold PBS and the events were acquired by FACSCalibur (BD Pharmingen, Franklin Lakes, NJ, USA). The collected data were analysed with WinMDI 2.9 software (Joseph Trotter, The Scripps Institute, La Jolla, CA, USA) after gating for the designated population.

The morphology, cell cycle and immunophynotype of murine compact bone-derived MSCs. A: Murine compact bone-derived MSCs exibit comparatively homogenious morphology. Bar=50 μm. B: The cell cycle phases of MSCs. C: Expression status of MHC-I and MHC-II molecules on MSCs.
The absolute and relative number of CD3+ cells in the spleens. At various time points post transplantation, the absolute number of nucleated splenocytes per spleen of the aGvHD and aGvHD+MSC groups of mice was determined. The relative number of CD3+ cells in the lymphocytes was gained by FCM results. The absolute number of CD3+ cells per spleen was obtained by multiplying the absolute number of nucleated splenocytes per spleen, the percentage of lymphocytes in nucleated splenocytes, and the percentage of CD3+ cell in the lymphocytes (relative number of CD3+ cells).
Syno-splenocyte infusion model. To exclude the non-specific injury effect of irradiation and/or injection, the syno-splenocyte infusion model was adopted. Nucleated splenocytes from female BALB/c mice were prepared after RBC lysis with 0.84% NH4Cl and an inoculum of 7×107 viable cells without (syno-splenocyte group) or together with 1×106 MSCs (syno-splenocyte+MSC group) in a total volume of 0.25 ml PBS was intravenously injected into sublethally irradiated (5 Gy) BALB/c mice (n=12). Specimens from at least three recipient mice per group were collected for experiments.
Statistical analysis. Results obtained from multiple repeat experiments were reported as the mean±standard error of the mean. Significance levels were determined by two-sided Student's t-test analysis. A probability (p) value <0.05 was considered statistically significant.
Results
Compact bone-derived murine MSCs. Compact bone-derived murine MSCs were of comparatively homogeneous morphology. The FCM analysis showed that the majority of the MSCs were in G0/G1 cell cycle phage (73.26%), a minor portion were in G2/M cell cycle phage (15.25%) and few cells were in apoptosis (0.59%). They constitutively expressed a medium amount of MHC-I molecules, but not MHC-II molecules (Figure1).

MSC infusion down-regulated the expression of CD1a and CD69 molecules on CD11b+ cells. A: The ratios of CD1a+CD11b+/CD11b+ of splenocytes in aGvHD and aGvHD+MSC mice at various time points after infusion. B: The ratios of CD69+CD11b+/CD11b+ of splenocytes in aGvHD and aGvHD+MSC mice at various time points after infusion. C, D: Representative results of A and B respectively, from three independent repeat experiments. *p<0.05, **p<0.01; D3: day 3, D5: day 5.
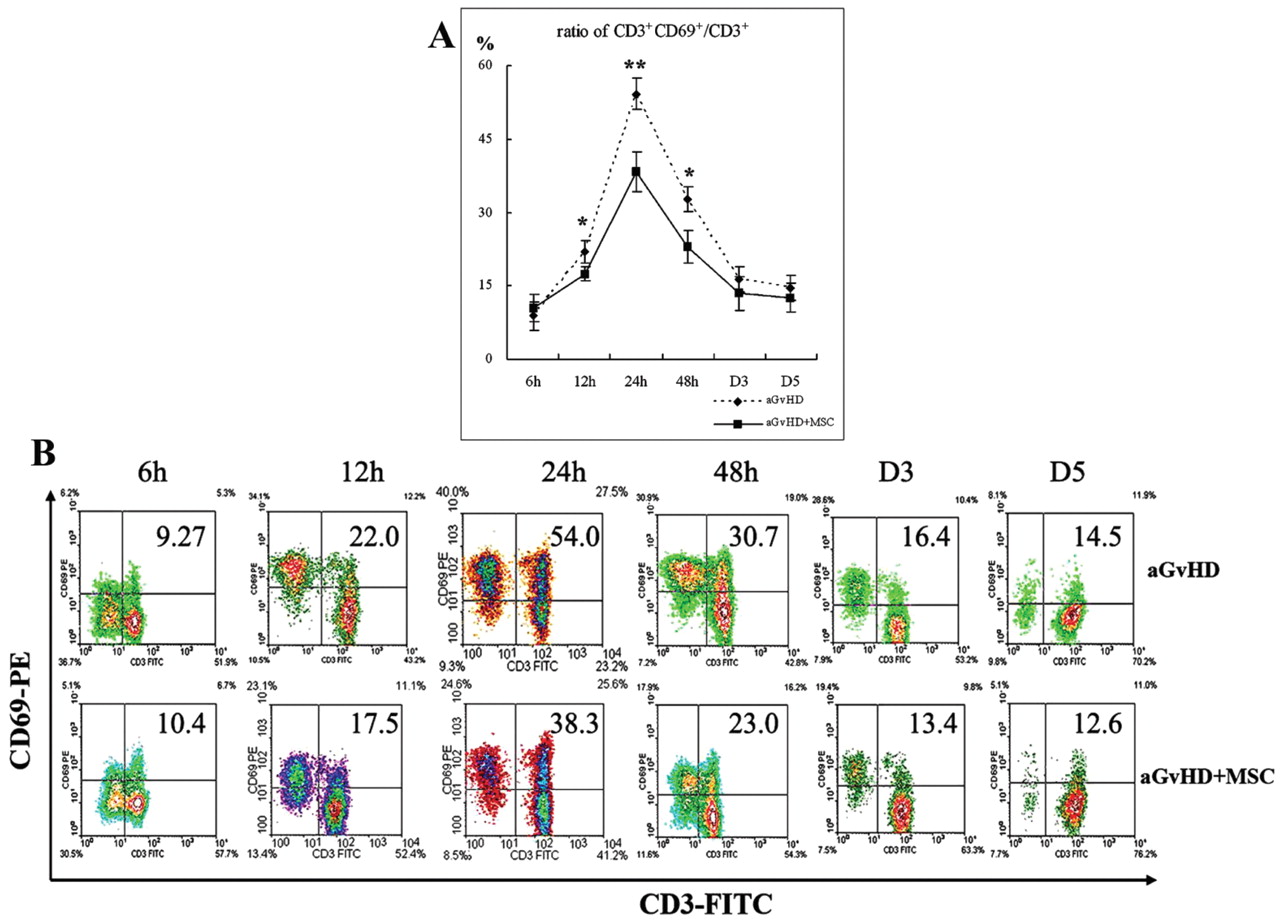
MSC infusion up-regulated the number and down-regulated the activation status of splenic CD3+ cells in aGvHD mice. A: The ratios of CD69+ CD3+/CD3+ cells of splenocytes in aGvHD and aGvHD+MSC mice at various time points after infusion. B: Representative result of A from three independent repeat experiments. *p<0.05, **p<0.01, D3: day 3, D5: day 5.
Down-regulated expression of MHC-II and CD69 molecules on splenic CD11b+ cells by MSC infusion. Antigen-presenting cells (APCs) are initiators of immune response both in vitro and in vivo. Macrophages which express the CD11b molecule on the cell surface are one kind of professional APC. The expression status of the MHC-II molecule and the early activation molecule CD69 were examined on splenic CD11b+ cells at various time points post transplantation. The results revealed that the ratios of MHC-II+CD11b+ to CD11b+ cells were significantly decreased at and after 12 hours; the ratios of CD69+CD11b+ to CD11b+ were also suppressed at 12, 24 and 48 hours after transplantation by co-infusion of MSCs (Figure 2A and B). Representative results of three repeat experiments are shown in Figures 2C and D.
Down-regulated splenic CD3+ cell activation by MSC infusion. Naïve T-cells activate rapidly on exposure to the co-stimulatory molecules and the antigen presented by APCs then generates armed effector T-cells (23). The in vivo immunoregulatory effects of MSCs were observed on T lymphocytes. Figure 3A shows the expression status of the early activation molecule CD69 on CD3+ cells at various time points post transplantation. The ratios were clearly reduced at 12 (23.45±2.33% vs. 16.57±1.85% for aGvHD and aGvHD+MSCs mice, respectively), 24 (57.38±3.96% vs. 36.57±3.85%) and 48 hours (33.48±2.69% vs. 22.77±3.33%) after infusion. Figure 3B shows representative results of three repeat experiments. The down-regulated activation status of T lymphocytes may have resulted in decreased effector T-cell formation and then decreased emigration from the spleen to aGvHD target tissues.
Up-regulation of number of splenic CD3+ cells by MSC infusion. At various time points post transfusion, the absolute and relative numbers of CD3+ cells per spleen of the two groups of mice were examined. As shown in Figures 4A and B, MSC infusion up-regulated the absolute and relative numbers of splenic CD3+ cells of aGvHD mice.
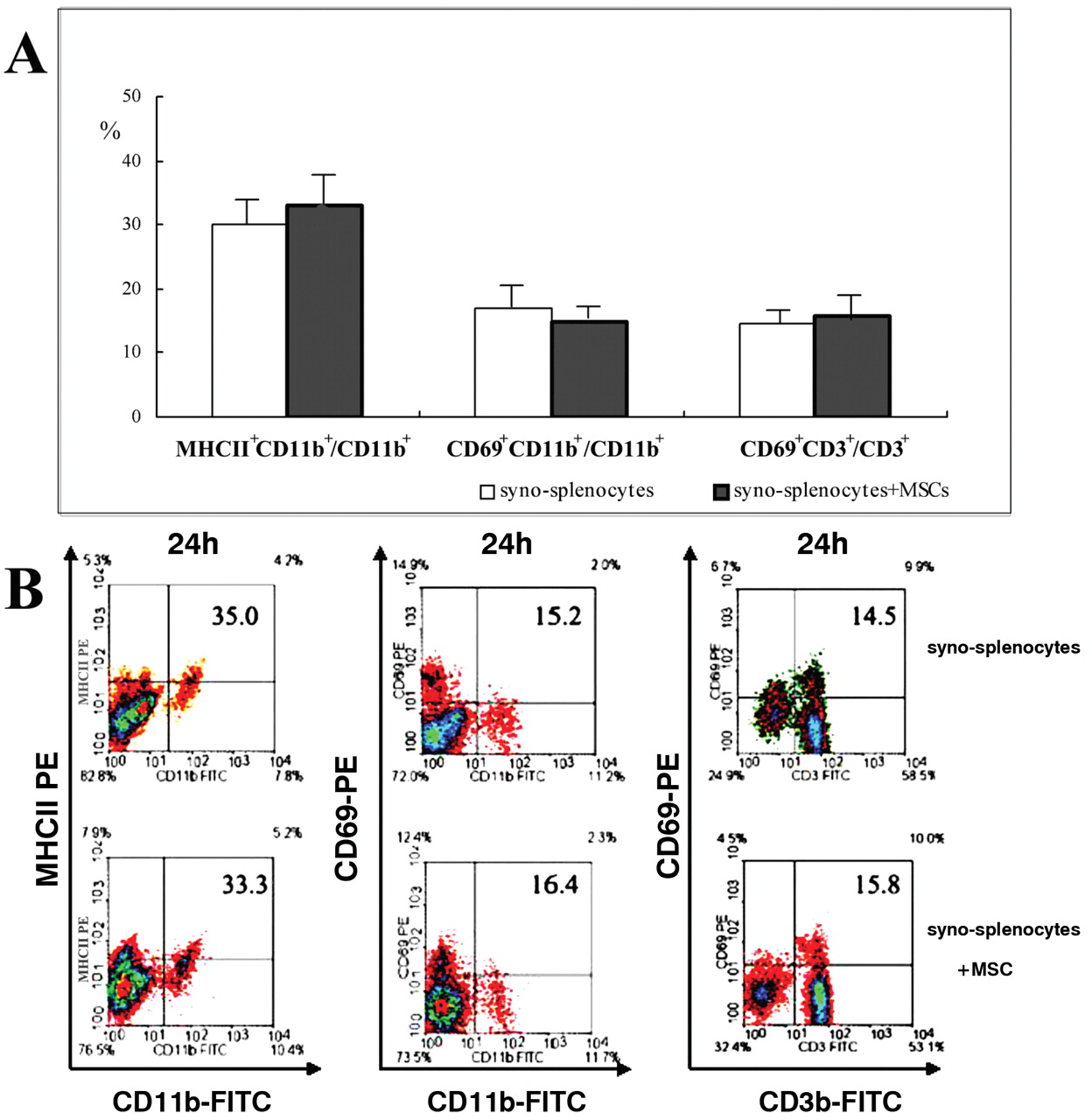
Activation situation in the syno-splenocyte infusion model. In order to exclude the non-specific activation of irradiation or infusion, the activation situation in the syno-splenocyte infusion model was examined at 24 hours after infusion, when there is a maximum activation effect in the aGvHD model. As shown in Figure 5A, the ratios of MHC-II+ CD11b+/CD11b+, CD69+CD11b+/CD11b+ and CD69+CD3+/CD3+ were not significantly affected. Representative results of three repeat experiments are shown in Figure 5B.
Discussion
MSCs are generally defined as a cell population that has the potential to differentiate along multiple lineages such as chondrocytes, osteocytes and adipocytes and they exist in bone marrow, adipose tissue, compact bone, placenta and other connective tissues (1-5). Recent data have also suggested that MSCs are characterised by their powerful immunosuppressive activity both in vitro (12) and in vivo (5, 16, 17-21), enhancing their therapeutic appeal for the prevention and management of aGvHD. However, the mechanisms underlying the immunomodulatory activities that MSCs play in vivo have not yet been well established.
Recently, it was reported that the murine compact bone is a good source from which to generate MSCs (5). Compact bone derived MSCs were used in the present study. The phenotypic features of splenocytes were investigated in a murine aGvHD model, which was created by sublethally irradiated BALB/c mice receiving splenocytes from an MHC incompatible strain, with or without co-infusion of cultured MSCs. In this model, previous studies have shown that MSCs infusion can delay aGvHD lethality (MSTs were 5.31±0.45 days vs. 7.89±0.83 days), ameliorate the pathological damage of the target tissues in aGvHD mice (24) and the transplanted MSCs can be detected in various tissues (25). All these findings provide supportive evidence that this model is suitable for the evaluation of the in vivo immunoregulatory activities of MSCs.
It is by now generally accepted that aGvHD can be summarised in a three-step process (26). Phase I is defined as the effect of conditioning, of which irradiation and chemotherapy injuries to host epithelium and endothelial cause APC maturation. Acting as one of professional APCs, macrophages (CD11b+ cells) will mature in the presence of inflammatory cytokines, enhancing the expression of the MHC-II molecule and the activation molecule CD69 in the phase I. CD69 antigen acts as a potent trigger of macrophage activation (27, 28). Activated macrophage expressing MHC-II molecules, which can present antigen peptides to T-cells, results in T-cell activation (29). Therefore, this study examined the expression status of the MHC-II molecule and the activation molecule CD69 on CD11b+ cells at various time points post transplantation. The result revealed that the ratios of MHC-II+ CD11b+/CD11b+ cells and the ratios of CD69+CD11b+/CD11b+ in the spleen of aGvHD mice were significantly decreased by MSC co-infusion (Figure 1A and B).

MSC infusion up-regulated the number of splenic CD3+ cells in aGvHD mice. A: The absolute number of splenic CD3+ cells of aGvHD and aGvHD+MSC mice at various time points after infusion. B: The relative number of splenic CD3+ cells of aGvHD and aGvHD+MSC mice at various time points after infusion as obtained by FCM analysis. *p<0.05, **p<0.01; D3: day 3, D5: day 5.

Activation situation of CD11b+ and CD3+ cells at 24 hours after infusion in the syno-splenocyte infusion model, when there was maxium activation in the aGvHD model. A: The ratios of CD1a+CD11b+/CD11b, CD69+CD11b+/CD11b and CD69+CD3+/CD3+ of splenocytes. B: Representative result of A from three independent repeat experiments. Syno-splenocyte and syno-splenocytes+MSC represent the BALB/c mice which received 7×107 BALB/c splenic nucleocytes without or with 1×106 MSC co-transfer, respectively.
During phase II of aGvHD, activated APCs and inflammatory cytokines induce donor T-cell activation and expansion. Naïve T-cells activate rapidly on exposure to the co-stimulatory molecules and host Ag presented by APCs, then proliferate and differentiate into effector T-cells. This Ag-driven proliferation and differentiation process is associated with the altered expression of several cell surface markers. The study of Wabnitz et al. showed that the co-stimulation-induced phosphorylation of L-plastin facilitates the surface transport of the T-cell activation molecules CD69 (30). Once expressed, CD69 also acts as a co-stimulatory molecule for T-cell activation and proliferation (31-33). Naïve T-cells activate rapidly on exposure to the co-stimulatory molecules and host Ag presented by APCs, and then generate armed effectors. In vitro investigations showed that the MSC-mediated inhibition of alloreactive T lymphocytes is associated with the down-regulation of the activation markers CD25, CD38, and CD69 detected both in CD4+ and CD8+ T lymphocytes (34). On the contrary, Ramasamy et al. reported that the expression of early activation markers of T-cells, namely CD25 and CD69, are not significantly altered by MSCs at 24, 48 and 72 hours in MLR (35). In the present study, the in vivo immunoregulatory effects of MSCs on T lymphocytes were observed. In the experimental murine aGvHD model, the ratios of CD69+CD3+/CD3+ cells in the spleen of aGvHD mice, which led to T lymphocyte activation, were suppressed by MSC co-transplantation (Figure 3A and B).
Finally, during phase III of aGvHD, effector T-cells migrate to the target organ and cause further injuries. CD69, acting as an early marker of T-cell activation, distinguishes an activated effector from naïve T-cells (36). Effector T-cells eventually egress from the second lymphocyte organ and migrate to the aGvHD relatively specific target organs, such as lung, gut, liver and skin, resulting in severe inflammation (37). Several clinical observations have shown that the pathologic lymphocytic infiltrates of target organs commonly occur during the period of profound lymphopenia (38). In the present study, the number of CD3+ cells per spleen was examined at various time points post transfusion. As shown in Figure 4, MSC infusion up-regulated the absolute and relative number of CD3+ cells in the spleens. The down-regulated activation status of T lymphocytes resulted in decreased effector T-cell formation. Therefore, a decrease in effector splenocyte migration from the spleen to the target organs may account for the increased number of CD3+ cells in the spleen and the reduced lymphocytic inflammation in the target organs.
In conclusion, this study demonstrated that MSCs can inhibit the three development stages of aGvHD. This study provided new clues for further detailed investigations needed to clarify the exact mechanisms underlying the modulation of MSCs on the ongoing immune response in vivo. Addressing this issue may contribute to the applications of MSCs in the clinical setting as a cellular therapy.
Acknowledgements
This study was supported by the National Key Basic Research Program of China (Grant number 2005CB522705, 2010CB833604), the National High Technology Research and Development Program of China (2007AA021109), the National Natural Sciences foundation of China (grant no. 30900568, 30730043, 30971068 and 30871018) and a Chinese High-tech Grant (no. 2007AA02Z454).
Footnotes
-
↵* These Authors contributed equally to this study.
- Received June 3, 2010.
- Revision received June 28, 2010.
- Accepted July 2, 2010.
- Copyright © 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
Reference
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.