


Abstract
Background: Therapeutic options in patients with metastatic colorectal cancer are still limited. As apoptosis contributes to the overall sensitivity to radiotherapy or chemotherapy, a better understanding of the apoptotic process in metastatic tumour tissues is necessary. Materials and Methods: Precision cut tissue slices (PCTS) of three human liver metastases were used to investigate the effect of activating CD95 antibodies (concentrations: 0.1 μg/ml, 1 μg/ml and 1 μg/ml and 1 μg/ml actinomycin D) as well as TNFα (concentrations 1 ng/ml; 10 ng/ml and 10 ng/ml and 1 μg/ml actinomycin D) directly in tumour tissue after 6 h, 12 h and 24 h. The apoptotic effect was assessed immunohistochemically. Results: Activating CD95 antibodies combined with actino-mycin D led to a significant increase in apoptosis after 12 h. Using TNFα at a high dosage, a significant increase in the apoptosis rate was observed after 6 h and after 12 h in all dosage groups. Conclusions: PCTS can be used to investigate the effect of different apoptotic signals directly in human tumour tissues. TNFα is able to effectively induce apoptosis in liver metastases of colorectal carcinoma. Thus, the extrinsic pathway of apoptosis may be a promising target in the development of new therapeutic approaches.
The liver is one of the organs most frequently involved in haematogenic metastatic spread of solid tumours. Primary tumour entities are, in particular, colon, gastric and breast carcinomas, but also melanomas and neuroendocrine tumours (1). A total of 15 to 25% of patients with colorectal cancer suffer from liver metastases by the time the primary tumour is diagnosed. An additional 35%-45% of patients will develop hepatic secondary tumours during the disease course (2, 3). Hepatic resection is the only curative treatment for liver metastases of colorectal cancer. Unfortunately, most liver metastases are, however, inoperable (4, 5). Further insight into the pathways of liver metastasis is necessary to develop adjuvant therapeutic strategies.
Experimental liver research can be performed using isolated primary liver cells, cell lines, whole liver models, artificial liver systems or precision-cut tissue slices (PCTS). Primary hepatocytes are used in drug development, for toxicity studies and in metabolic and virological studies (6). PCTS have been used in pharmacology and toxicology in a variety of studies (6). The method of liver PCTS has been adapted for functional morphological research in liver tissue, namely for apoptosis investigation (1).
Cell death is considered to comprise several types of cellular demise including apoptosis, autophagy, necrosis and mitotic catastrophe (7, 8). Apoptosis was first mentioned by Kerr et al. who described a main morphological form of cell death characterised by a reduction of cell volume, chromatin condensation, nucleus fragmentation and plasma membrane maintenance until late in the cell death process (9, 10). Two distinct signalling pathways regulate apoptosis, the mitochondrial or intrinsic pathway and the extrinsic pathway via cell death receptors (11). In the extrinsic pathway, natural killer cells or cytotoxic T-cells produce pro-apoptotic ligands which belong to the tumour necrosis factor (TNF) family (12). These activate pro-apoptotic death receptors on the surface of the target cells (13, 14). Both pathways stimulate pro-apoptotic members of the cysteine protease family of caspases (15, 16). The active effector caspases cleave cell proteins and lead to DNA fragmentation (10).
In this study PCTS of human liver metastases were used to investigate the effect of activating the extrinsic pathway by CD95 as well as by TNFα directly in the tumour tissue, and the apoptotic effects were assessed.
Rate of apoptosis per 10 HPF after stimulation with an activating CD95 antibody in concentrations of 0.1 μg/ml, 1 μg/ml and 1 μg/ml with actinomycin D 1 μg/ml as well as TNFα in concentrations of 1 ng/ml, 10 ng/ml and 10 ng/ml with actinomycin D 1 μg/ml after 6 h, 12 h and 24 h. Data are given as mean value (±standard error of the mean).
Material and Methods
Patient data. For the investigation of CD95-induced apoptosis, PCTS of three resection specimens of liver metastases of colorectal carcinoma were used. The donors were two female and one male patient with a mean age of 66 years. The mean diameter of the liver metastases was 6.0 cm. All three cases showed moderate histological grading. TNFα studies were performed with tissue from resected liver metastases from two male and one female patient with a mean age of 65.7 years. All tumours showed moderate grading. The mean diameter of metastases was 5.3 cm.
The investigations were approved by the Ethical Committee of the University of Cologne. The patients gave their informed consent for the study.
PCTS. The slices were prepared as described elsewhere (1). In brief, cylindrical tissue cores with a diameter of 0.8 cm were removed from the metastatic tissue. From these cores, 200-300 μm thick slices were cut using the Brendel Vitron tissue slicer (Vitron, Tuscon, AZ, USA), filled with ice-cold oxygenated physiological Ringer's solution. Incubation was performed in a static incubation system using 34-well plastic tissue culture plates (VWR, Darmstadt, Germany) on a rocker platform (Heidolph, Schwabach, Germany) at 37°C with saturated humidity and 5% CO2 in 40 ml RPMI medium, supplemented with 0.5 ml penicilline/streptomycine, 0.48 ml l-glutamine, 2.4 ml horse serum, 8 ml foetal calf serum and 0.24 ITS supplement (all supplied by SIGMA, Taufkirchen, Germany). After an equilibration time of 30 min, a medium change was performed. Following a resting time of 3 h, stimulation with the activating CD95 antibody EOS9.1 (Becton Dickinson, Pharmingen, San Diego, CA, USA) in concentrations of 0.1 μg/ml, 1 μg/ml and 1 μg/ml with addition of 1 μg/ml actinomycin D (Lyovac; Merck, Harleem, the Netherlands) was performed. TNFα was obtained from TEBU, Offenbach, Germany and used for stimulation at the following concentrations: 1 ng/ml, 10 ng/ml and 10 ng/ml with 1 μg/ml actinomycin D. Appropriate negative controls were performed.
After 6 h, 12 h and 24 h, the slices were removed, fixed in 4% neutral buffered formalin overnight and processed routinely for histological and immunohistological investigations.
Processing of the slices. From the paraffin-embedded tissues, 4 μm thick slices were cut for conventional histology (H&E staining). Apoptosis was assessed immunohistochemically using M30 antibody (CytoDeath; Roche, Mannheim, Germany). In brief, after deparafinisation and blocking of endogenous peroxidase, the primary antibody M30 was incubated overnight at 4°C at a dilution of 1:50. Detection was performed using monoclonal secondary biotinylated rabbit anti-mouse antibody 1:200 (DAKO, Glostrup, Denmark) followed by a streptavidin-biotin-complex (Vector, Burlingame, CA, USA) and visualised with DAB (DAKO). For evaluation, positive signals in 10 high power fields (HPF) were counted using an ocular grid.

Rate of apoptotic events after stimulation with CD95. Data represent mean values and bars represent the standard error of the mean. AD: Actinomycin D; LK: negative control.
Statistical analysis. For statistical analysis, SPSS for Windows (now IBM, Chicago, IL, USA) was used. The Mann-Whitney U-test and Student's t-test were performed for non-parametrical and parametrical data analysis, respectively. A p-value <0.05 was considered to be statistically significant. Statistical analysis was supported by Mr. Hahn of the Department of Medical Statistics, University of Cologne.
Results
CD95-induced apoptosis. Stimulation of apoptosis was observed with evenly distribution of apoptotic cells in the metastatic tissue (Table I and Figures 1 and 2).
After 6 h of stimulation, an increase in the apoptotic rate was observed in the group treated with 0.1 μg/ml (2.1-fold), 10 μg/ml (4-fold) and 10 μg/ml CD95 with 1 μg/ml actinomycin D (3.25-fold). These results, however, were not statistically significant (p=0.063).
After 12 h, the apoptotic rate was higher in the groups treated with 0.1 μg/ml CD95 (2.3-fold) and 1 μg/ml (2.6-fold). These results were not statistically significant. Adding 1 μg/ml actinomycin D to the group treated with 1 μg/ml CD95 led to a 4-fold increased apoptosis which was statistically significant (p=0.001).
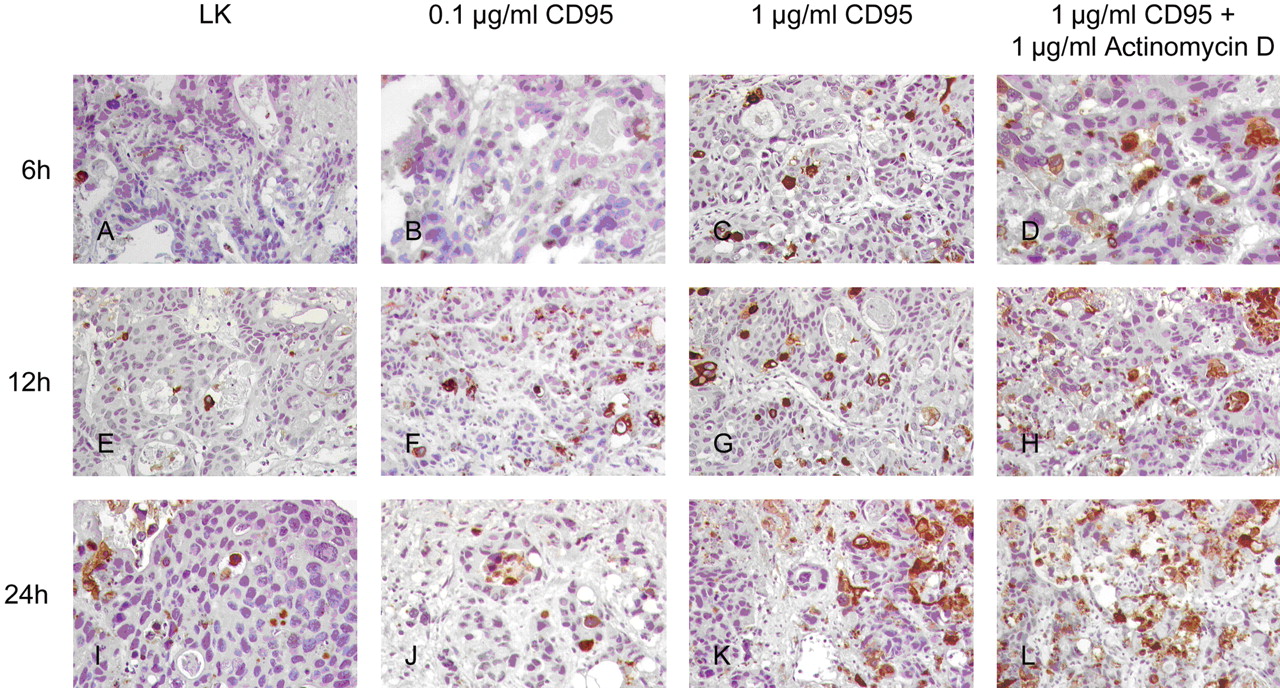
Immunohistological detection of apoptotic events using M30 Cytodeath after stimulation of the CD95 system. Low dose of the activating antibody did not induce an increased rate of apoptosis (B, F, J) compared to a higher dose (C, G, K). Adding actinomycin D increased the apoptotic rate (D, H, L). LK: Negative control.
After 24 h, the stimulating effect of the activating CD95 antibody was reduced, with a 1.3-fold increase in apoptotic events in the treatment group treated with 0.1 μg/ml CD95 and a 2.0-fold increase in the group treated with 1 μg/ml CD95. Adding actimomycin D revealed a 2.5-fold increase in apoptotic events which was statistically significant (p=0.006).
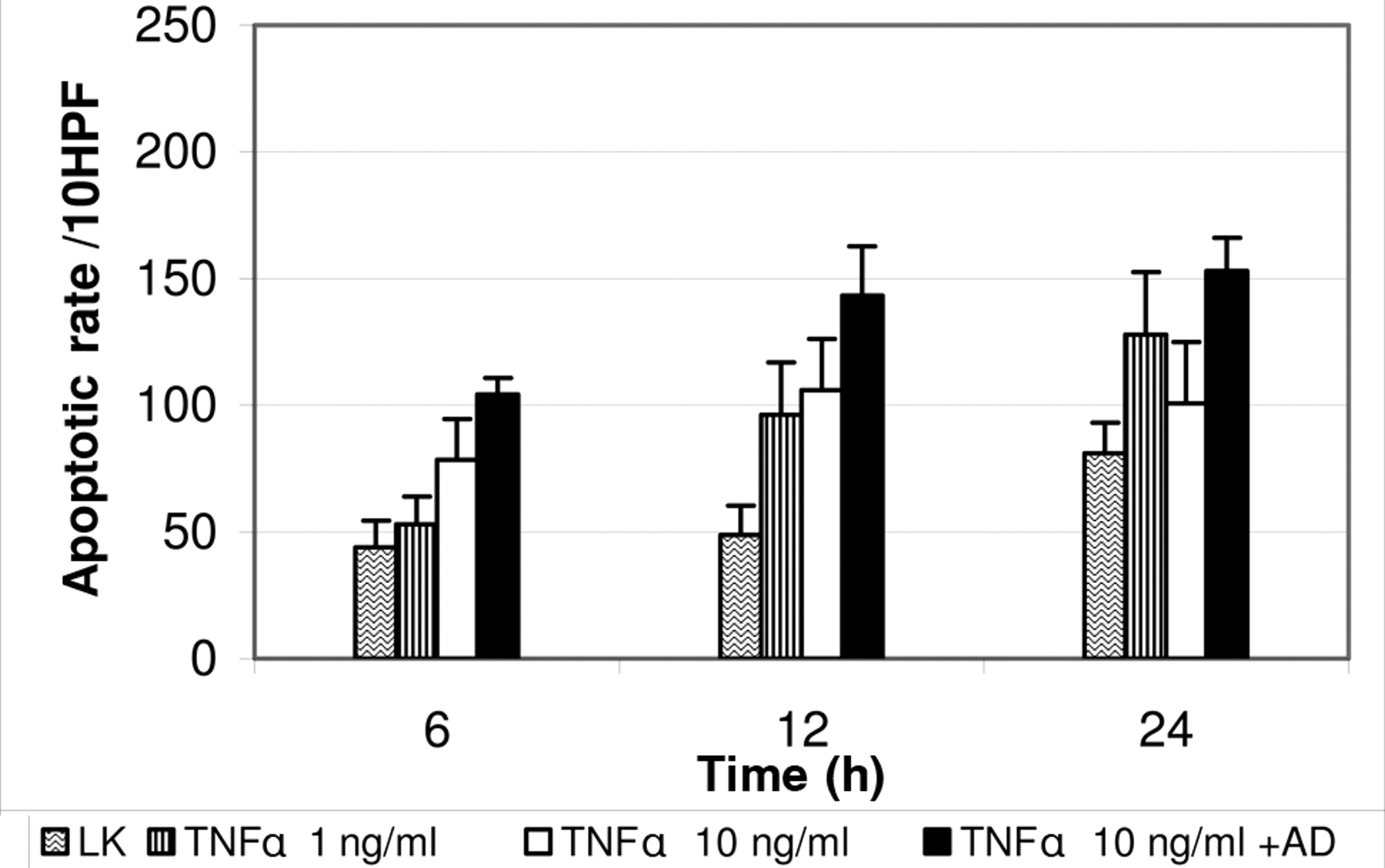
TNFα-induced apoptosis. After 6 h, PCTS of liver metastases incubated with TNFα showed an increase in the apoptotic rate depending on the concentration (Figures 3 and 4). A slight increase in the apoptotic rate was seen in the PCTS treated with a low dose of TNFα (1 ng/ml). Using higher doses of TNFα (10 ng/ml), a significant increase by a factor of approximately 1.8 was observed (p=0.04). TNFα at high concentration and actinomycin D yielded a significant 2.4-fold increase in apoptotic events (p=0.013).
After 12 h, all groups showed a significant increase in the apoptotic rate compared to spontaneous apoptotic events, with a 2.0-fold increase after stimulation with 1 ng/ml TNFα (p=0.008), a 2.2-fold increase with 10 ng/ml TNFα (p=0.031) and a 3-fold increase after stimulation with 10 ng/ml TNFα and 1 μg/ml actinomycin D (p=0.041).
The apoptotic effect was reduced after 24 h incubation. The slices treated with 1 ng/ml TNFα showed a 1.6-fold increase of apoptosis, the slices treated with 10 ng/ml TNFα showed a 1.2-fold increase and the slices treated with 10 ng/ml TNFα and 1 μg/ml actinomycin D showed a 1.8-fold increase in apoptosis rate. The results after 24 h were not statistically significant (p>0.05).

Rate of apoptotic events after stimulation with TNF alpha. Boxes represent mean value and bars represent standard error of the mean. AD: Actinomycin D; LK: negative control.
Discussion
Current in vivo models, for example in pigs, rats, rabbits and mice, are difficult to standardise and do not represent all human pathomechanisms. In addition, modern ethical requirements demand the reduction of the use of animals for scientific purposes. Therefore, in this study, the PCTS method was adopted for morphological investigations (17).

Apoptosis after stimulation with TNFα. After 6 h, the slices treated with 10 ng/ml TNFα showed an increased apoptotic rate (C, D), whereas the slices treated with the low dose of 1 ng/ml were mainly unstained. After 12 h, however, in all groups, an increased rate of cell death was seen (F, G, H). This was not further increased after 24 h (J, K, L). LK: Negative control.
The PCTS method has several advantages compared to other models. Preparation of PCTS does not require the use of proteolytic enzymes. The slices retain the original architecture. Thus, cell-to-cell and cell-to-matrix interactions are preserved. Compared to in vivo models, PCTS allows the direct and rapid analysis of tissue under standardised and lifelike conditions in a reproducible manner (1, 6).
Failure to eliminate cancer cells that have been exposed to cytotoxic drugs may contribute to the development of tumour resistance (18). Apoptosis is considered to contribute to the overall sensitivity to radiotherapy or chemotherapy. Cells that are able to avoid apoptosis will be resistant to such therapy (19). Apoptosis can be induced via the mitochondrial (intrinsic) pathway or via extrinsic signalling. Intracellular events such as DNA damage and several chemotherapeutica drive apoptosis mainly through the intrinsic pathway (11). In metastases, as a late stage of tumour progression, the extrinsic pathway seems to be a promising therapeutic target (20, 21). Apoptotic cell death activation as anticancer approach has the potential to induce tumour regression rather than to stop tumour growth (10). Several targets for activation of the extrinsic pathway of apoptosis as a therapeutic strategy are under investigation (20).
In this study, the investigation was focused on the extrinsic pathway analysing apoptosis induced via CD95 receptor and TNFα. Using PCTS, it was shown that effective induction of apoptosis is possible in human tissue with preserved architecture. It is a dose- and time-dependent process. For cell lines of colorectal carcinoma, a cytotoxic effect of TNFα has been demonstrated (22). Using TNFα at high concentration as an inductor of apoptosis, a significant cell death rate was observed already after 6 h in our study. After 12 h, the effect was seen in all concentration groups. These observations demonstrate that the extrinsic pathway via TNFα is an effective way to induce apoptosis in all investigated liver meta-stases. In the literature, several resistance mechanisms for TNFα-induced apoptosis have been described. They include up-regulation of transcription factors such as nuclear factor kappa B (23-25). The present study did not further investigate whether these factors play an important role.
The reduced effect after 24 h may have different causes. Firstly, apoptosis is a short-term reaction of cells (10). Cells sensitive to apoptosis induction will be destroyed after a short period of time. Secondly, it is unclear, whether the concentration of TNFα used led to a saturated solution; thus, complete consumption cannot be excluded. Thirdly, the initial deterioration process of the slices could discharge proteins interacting non-specifically with the triggering molecule.
In an earlier study, liver metastases of colorectal carcinomas were shown to induce a dense lymphocytic infiltrate (26). Tumour-infiltrating lymphocytes are known to possess Fas ligand on their surface as one effector mechanism of cytotoxicity. In the present study, this pathway was investigated directly in human tissue of liver metastases. Using CD95-activating antibodies, a high dosage was necessary to induce a significant response after 12 h. After 24 h this effect was lost. CD95 seems to be less effective in induction of apoptosis in the system of PCTS. This could have biological causes. Tumour cells may lose the CD95 receptor on the surface of their cells as part of their survival strategy. A technical cause may also be possible. Since an activating antibody was used in this system and antibodies have a complex protein structure, non-specific binding to released proteins in the supernatant could reduce or abolish the effect.
In summary, PCTS can be used to investigate the effect of different apoptotic signals directly in human tumour tissues. TNFα is able to effectively induce apoptosis in liver metastases of colorectal carcinoma. Thus, the extrinsic pathway of apoptosis may be a promising target in the development of new therapeutic approaches.
Acknowledgements
This work is dedicated to Professor Dienes with gratitude for his support.
- Received June 6, 2010.
- Revision received June 28, 2010.
- Accepted July 6, 2010.
- Copyright © 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved
References
In this issue




{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.