


Abstract
Aim: Short-term oestrogen and progesterone treatment (STEPT) mimics the pregnancy hormone milieu. This study compared the development of N-methyl-N-nitrosourea (MNU)-induced mammary cancer in female Lewis rats that received STEPT in early or later life. Materials and Methods: Rats in Groups 1 and 2 received a single intraperitoneal injection of 50 mg/kg MNU at 4 weeks old. Pellets containing 0.5 mg 17β-estradiol and 32.5 mg progesterone (EP) were subcutaneously implanted in rats in Group 1 during 6-9 weeks old. Rats in Groups 3 and 4 received 50 mg/kg MNU at 22 weeks old and again at 23 weeks old. EP pellets were implanted in rats in Group 3 during 24-27 weeks old. At the time of EP removal and 8 weeks afterward, 4 randomly selected rats in each group were sacrificed for blood sampling. The fatty acid composition of serum phospholipids was measured by capillary gas chromatography. The remaining rats were sacrificed when they developed mammary tumours ≥1 cm in diameter or at the termination of the experiment, which was at 18 weeks old for Groups 1 and 2 and at 64 weeks old for Groups 3 and 4. Mammary cancer was histologically confirmed. Results: Group 1 had a significantly suppressed incidence of mammary cancer compared to Group 2 (7% vs. 90%), whereas the cancer incidence in Group 3 was similar to that of Group 4 (50% vs. 56%). Rats in Group 1 had significantly smaller n-6/n-3 polyunsaturated fatty acid (PUFA) ratios and higher levels of docosahexaenoic acid (DHA) than those in Group 2 at the time of EP removal but not 8 weeks after EP removal. Neither the PUFA ratios nor the DHA levels differed between Groups 3 and 4 at any time. These data suggest that the age at which STEPT is administered is important, since its mammary cancer-suppressing potential was lost in aged animals. Conclusion: DHA and the n-6/n-3 PUFA ratio may play a crucial role in mammary cancer suppression by STEPT.
Women who experience a full-term pregnancy at a young age have a substantially lower risk of breast cancer (1-4). For women who undergo a full-term pregnancy before the age of 20 years, the risk of developing breast cancer is one-half of the risk in nulliparous women. Although early full-term pregnancy provides long-lasting protection against breast cancer, parity protection becomes weaker with increasing age, and pregnancy after the age of 35 years appears to increase the breast cancer risk (1-5). Parity-induced protection against breast cancer occurs in women of all ethnicities and is the only normal physiological condition that consistently protects against breast cancer without known side-effects. Parity protection can be seen in the N-methyl-N-nitrosourea (MNU)-induced rat mammary cancer model (6). Young rats (≤3 months of age) that undergo a full-term pregnancy before, during or shortly after carcinogen exposure have a significantly reduced overall mammary cancer incidence and multiplicity and have a prolonged latency (reviews: 7-9). The pregnancy hormone milieu can be mimicked by oestrogen and progesterone (EP) treatment. Parity-induced protection against mammary cancer has been reproduced in rats by short-term EP treatment (STEPT) that lasts 21 days, which is approximately equivalent to the gestational period of rats (7-10). The benefits of substituting EPT for pregnancy are that the treatment duration can be easily controlled and that the effects can be studied for animals at any age. However, oestrogen is a potent mammary cancer-enhancing agent, and long-term EP exposure increases the risk of cancer. Therefore, the duration of exposure in studies of mammary cancer protection in rats should be <20 weeks (10). Moreover, late STEPT (LSTEPT) exposure in aged rats (MNU at 7 weeks of age and EPT during 24-27 weeks of age) accelerates mammary carcinogenesis compared with non-EPT animals (11).
The cellular and molecular mechanisms that underlie parity-induced protection against breast cancer are largely unknown. Local changes in the mammary gland and/or systemic alterations of the host environment appear to be involved. Rat studies have shown that parity promotes the differentiation of the mammary glandular tree and decreases the proliferation of mammary epithelial cells (12, 13). Rats who experience pregnancy or receive STEPT at a young age (early STEPT [ESTEPT]) exhibit characteristic changes in their mammary glands; specific genes involved in epithelial cell differentiation are upregulated, and genes involved in epithelial cell proliferation are down-regulated (13-16). Moreover, these rats are especially refractory to carcinogen-induced mammary epithelial cell proliferation (13, 17). When isolated mammary epithelial cells from MNU-treated virgin rats were transplanted into isogenic parous and age-matched control hosts, the rate of cancer development was lower in the parous rats than in the age-matched controls (18). These observations suggest that the resistance to mammary carcinogenesis in parous rats may be caused by persistent systemic changes in the host environment.
Epidemiological studies have suggested an association between the quality and the quantity of dietary fat and the development and progression of breast cancer. Although precise analysis of dietary fatty acids is limited in human studies, studies of experimental animals have yielded clear results. Tannenbaum and Silverstone first demonstrated that the amount of dietary fat enhances the development of chemically induced mammary cancer in experimental animals (19). Moreover, data from animal experiments indicate that the mammary cancer-promoting properties may be due to differences in the fatty acid composition (20, 21). In rodents, a high-fat diet rich in n-6 polyunsaturated fatty acid (PUFA), especially linoleic acid, stimulates mammary cancer growth and metastasis, whereas n-3 PUFA, such as eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) have a cancer-suppressing effect. The fatty acid composition of serum or plasma phospholipids reflects the medium-term (weeks to months) qualitative dietary intake of fatty acids (22). Therefore, levels of non-fasting serum phospholipids can serve as a reliable source of data (23). In humans, changes in the fatty acid composition of serum phospholipids during the course of pregnancy cannot be explained by changes in dietary intake alone (24, 25). Changes in the hormonal status may explain the differences in serum fatty acid composition. In rats, the expression of genes related to fatty acid synthesis, such as stearoyl coenzyme A desaturase 2 (Scd2) and fatty acid elongase 2 (rELO2) is altered in parous mammary glands compared to age-matched controls (13). In the present study, the aim was to determine the relationship between changes in the fatty acid composition of serum phospholipids and MNU-induced mammary carcinogenesis by studying rats that consumed the same diet and did or did not receive short-term exposure to a hormonal milieu mimicking pregnancy.
Materials and Methods
Animals. Three-week-old female Lewis rats were purchased from Charles River Japan (Atsugi, Japan). The animals were housed in a plastic cage with paper bedding (Paper Clean, SLC, Hamamatsu, Japan), 3-4 rats per cage, in a temperature-(22±2°C) and humidity- (60±10%) controlled animal room under a 12-h light/dark cycle. Rats were fed a commercial pellet diet (CMF 30 kGy; Oriental Yeast, Chiba, Japan) and water ad libitum throughout the experiment. All animal experiments were approved by the Animal Experimentation Committee of Kansai Medical University.
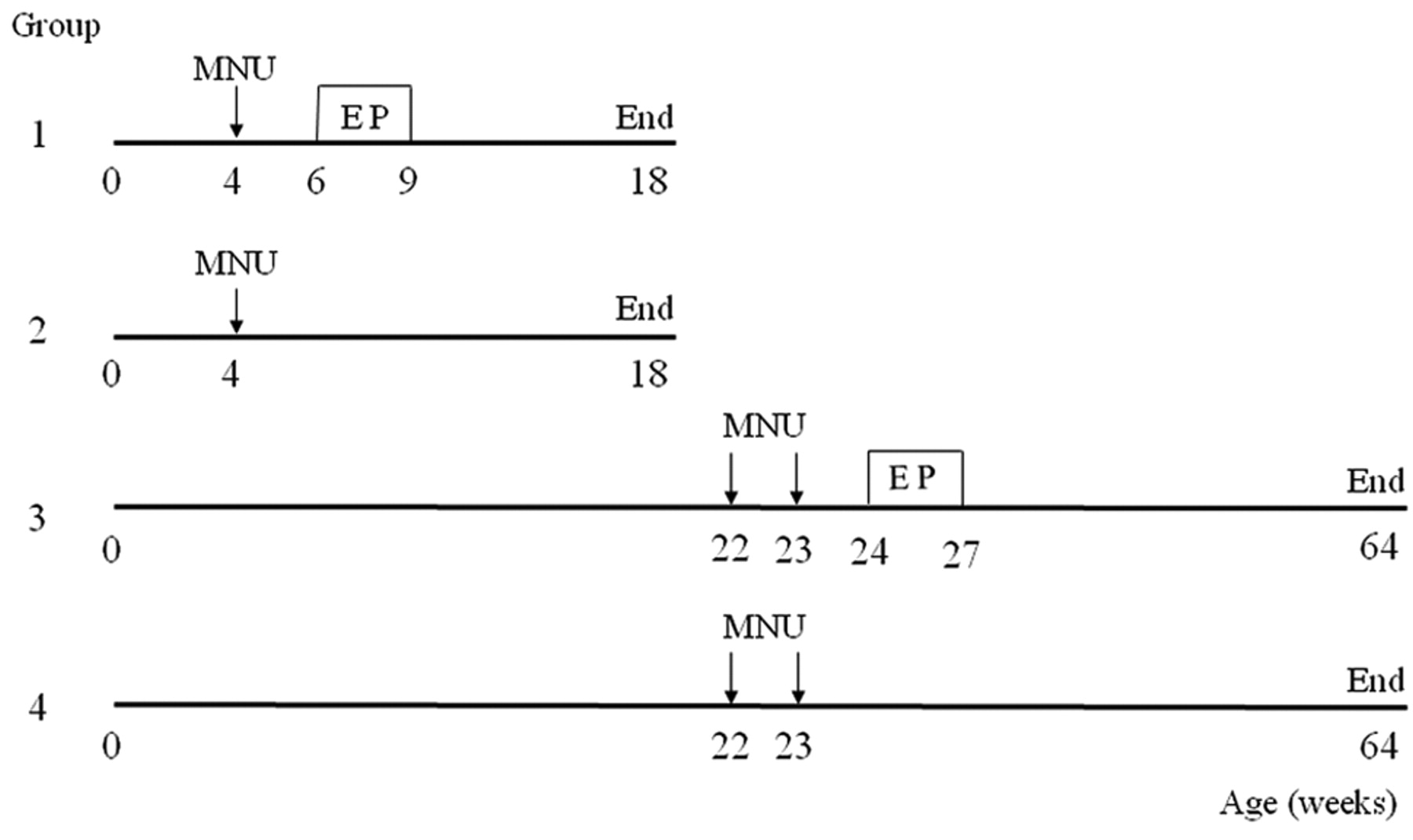
Experimental procedure. Rats were injected intraperitoneally with 50 mg/kg MNU (Sigma, St. Louis, MO, USA) at 4 weeks of age (Groups 1 and 2) or twice at 22 and 23 weeks of age (Groups 3 and 4). The MNU was stored at −20°C in the dark and dissolved in physiological saline containing 0.05% acetic acid immediately prior to injection. A steady-state, 21-day release, EP pellet containing 0.5 mg 17β-estradiol and 32.5 mg progesterone (Innovative Research of America, Sarasota, FL, USA) was implanted subcutaneously into the back at 6 weeks of age (Group 1) or 24 weeks of age (Group 3). The pellets were removed 3 weeks after implantation. The EP pellet significantly elevates circulating 17β-estradiol and progesterone levels, which reach pregnancy levels 2 weeks after implantation and decrease to control levels 8 weeks after implantation (10). The rats in Groups 2 and 4 did not receive EP pellets and comprised the age-matched controls. At the time of EP pellet insertion, all rats (including the age-matched controls) were checked for mammary tumours by palpation, and any tumour-bearing rats were excluded from the experiment. The experimental protocol is shown in Figure 1.
Mammary tumour detection and sacrifice. The rats were checked weekly by palpation for mammary tumours. Rats were sacrificed when their largest mammary tumour reached a diameter of ≥1 cm. Rats that did not develop a 1-cm mammary tumour were sacrificed at 18 weeks of age (Groups 1 and 2) or at 64 weeks of age (Groups 3 and 4), and the experiment was terminated.
Histological examination. Mammary tumours ≥1 cm in diameter and all six pairs of non-tumoural mammary glands were removed at autopsy and fixed in 10% neutral buffered formalin. All fixed tissues were embedded in paraffin, cut into 4-μm sections, and stained with hematoxylin and eosin (HE).

Schematic representation of the experimental protocols. Group 1 received early short-term oestrogen and progesterone treatment (ESTEPT), and Group 2 was its age-matched control. Group 3 received late short-term oestrogen and progesterone treatment (LSTEPT), and Group 4 was its age-matched control. MNU, N-Methyl-N-nitrosourea; EP, oestrogen and progesterone treatment.
Mammary carcinogenesis. One rat from Group 2 and two rats from Group 3 that were sacrificed due to the development of a mammary tumour that reached 1 cm in diameter were histologically diagnosed as having benign mammary tumours; therefore, these three rats were excluded from the study. All other mammary tumours ≥1 cm were histologically confirmed to be mammary cancer. The incidence of cancerous mammary tumours of ≥1 cm and the latency (interval from MNU treatment until sacrifice) were calculated. Additionally, the multiplicity (including histologically detected mammary tumors of any size) was compared.
Fatty acid analysis in serum phospholipids. To determine fatty acid composition, blood samples were collected from four randomly selected rats per group that were sacrificed at the time of EP pellet removal or 8 weeks later (when mammary glands were free of EP effects and presumed to be involuted). The animals were anaesthetized with diethyl ether, and blood was collected by cardiac puncture. Sera were separated from the non-fasting whole blood samples, centrifuged for 5 min at 3,000 rpm, and stored at −80°C until use. The fatty acid composition of the total phospholipid fractions of serum was determined. Total lipids were extracted by the method of Bligh and Dyer (26). The total phospholipid fraction was separated by thin-layer chromatography. For an internal standard, 1, 2-diheptadecanoyl-sn-glycero-3-phosphocholine (Avanti Polar Lipids, Inc., Alabaster, AL, USA) was added. Total phospholipid fractions were transmethylated with HCl-methanol, then the fatty acid composition was analysed by gas chromatography (GC-2014; Shimadzu Corporation, Kyoto, Japan) with a capillary column DB-225 (0.25 mm × 30 m × 0.25 μm; J&M Scientific, Folsom, CA, USA). The whole system was controlled with gas chromatography software (GC-Solution; Shimadzu Corporation). Fatty acid composition expressed as the area percentage of total fatty acids was compared as individual fatty acid, saturated fatty acid (SFA), monounsaturated fatty acid (MUFA), and PUFA. PUFA are fatty acids of 18 carbons or more in length with 2 or more double bonds. PUFA were divided into n-6 and n-3, depending on the position of the double bond relative to the methyl end of the fatty acid molecule, and the n-6/n-3 ratio was calculated.

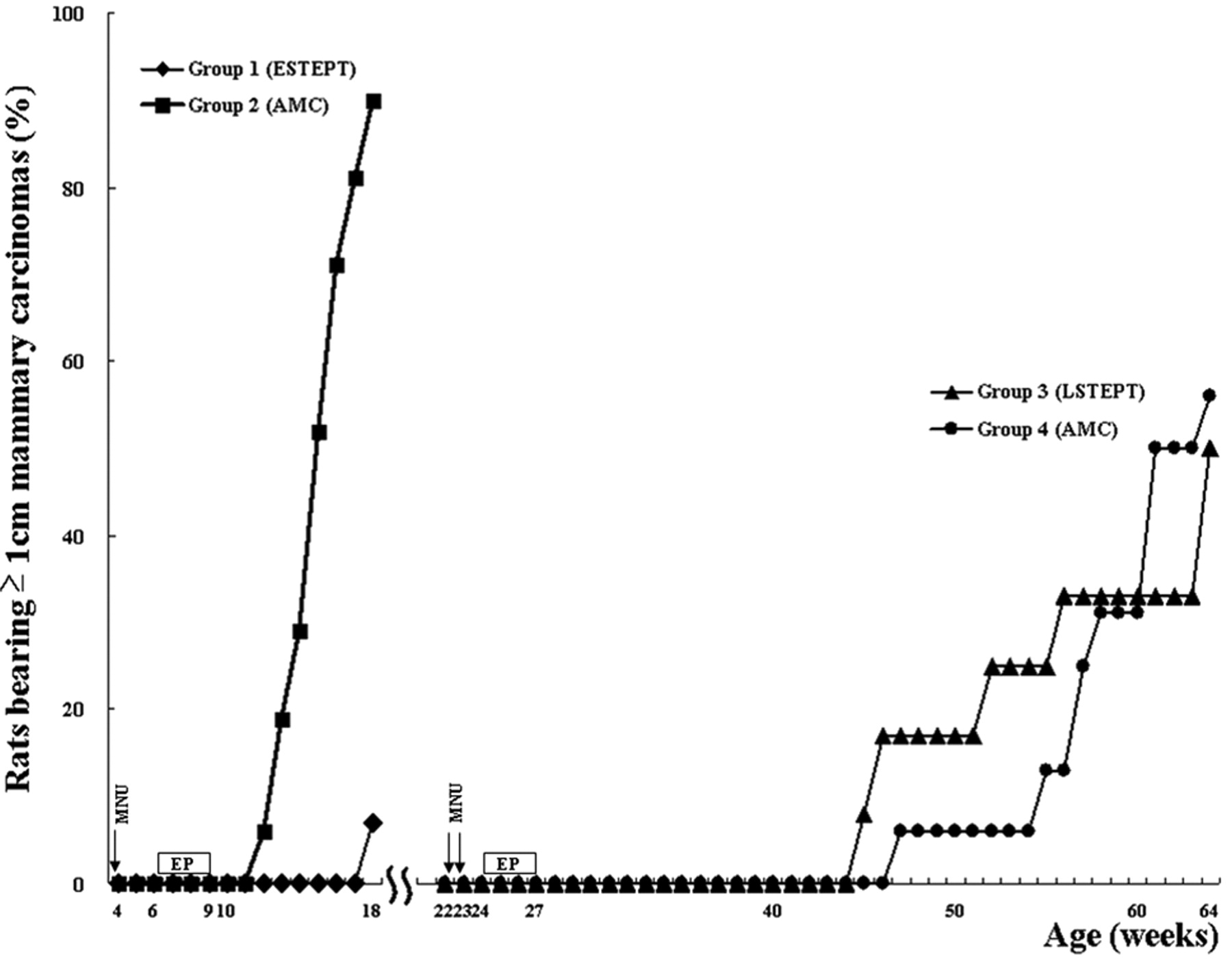
Effects of early, and late short-term oestrogen and progesterone treatment (ESTEPT or LSTEPT) on cumulative incidence of MNU-induced mammary cancers ≥1 cm in diameter in female Lewis rats. AMC, Age-matched controls.
Statistical analysis. All values were expressed as the mean±standard error (SE). Cancer multiplicity and latency and the levels of fatty acids were analyzed by Student's t-test, and the incidence of mammary cancer was analyzed using the χ2 test. A probability value of p<0.05 was considered statistically significant.
Results
Mammary carcinogenesis. Among the rats that received ESTEPT (Group 1), two rats (7%, 2/28) developed mammary cancer at the termination of the experiment when they were 18 weeks old (14 weeks after MNU). However, the age-matched controls (Group 2) started to develop mammary cancer (≥1 cm) from 12 weeks of age (8 weeks after MNU); the mammary cancer incidence in Group 2 gradually increased with time and reached 90% (28/31) at 18 weeks of age, and terminated the experiment (p<0.01, Figure 2). The latency was significantly longer in the ESTEPT group compared to the age-matched controls (14.0 vs. 11.1 weeks). The cancer multiplicity was significantly smaller in the ESTEPT group (0.3 vs. 3.7, Table I). Thus ESTEPT significantly suppressed the mammary cancer yield. In contrast, LSTEPT (Group 3) did not suppress the development of mammary cancer (≥1 cm) compared with age-matched controls (Group 4, Figure 2); the final mammary cancer incidence was 50% (6/12) in Group 3 and 56% (9/16) in Group 4. Although the cancer multiplicity was significantly smaller in LSTEPT compared with respective age-matched controls (0.7 vs. 1.9), there was no difference in latency (Table I). Thus, LSTEPT did not significantly suppress the mammary cancer yield as evaluated by mammary cancer incidence (Figure 2).
Effects of short-term oestrogen and progesterone treatment on MNU-induced mammary carcinogenesis in female Lewis rats.
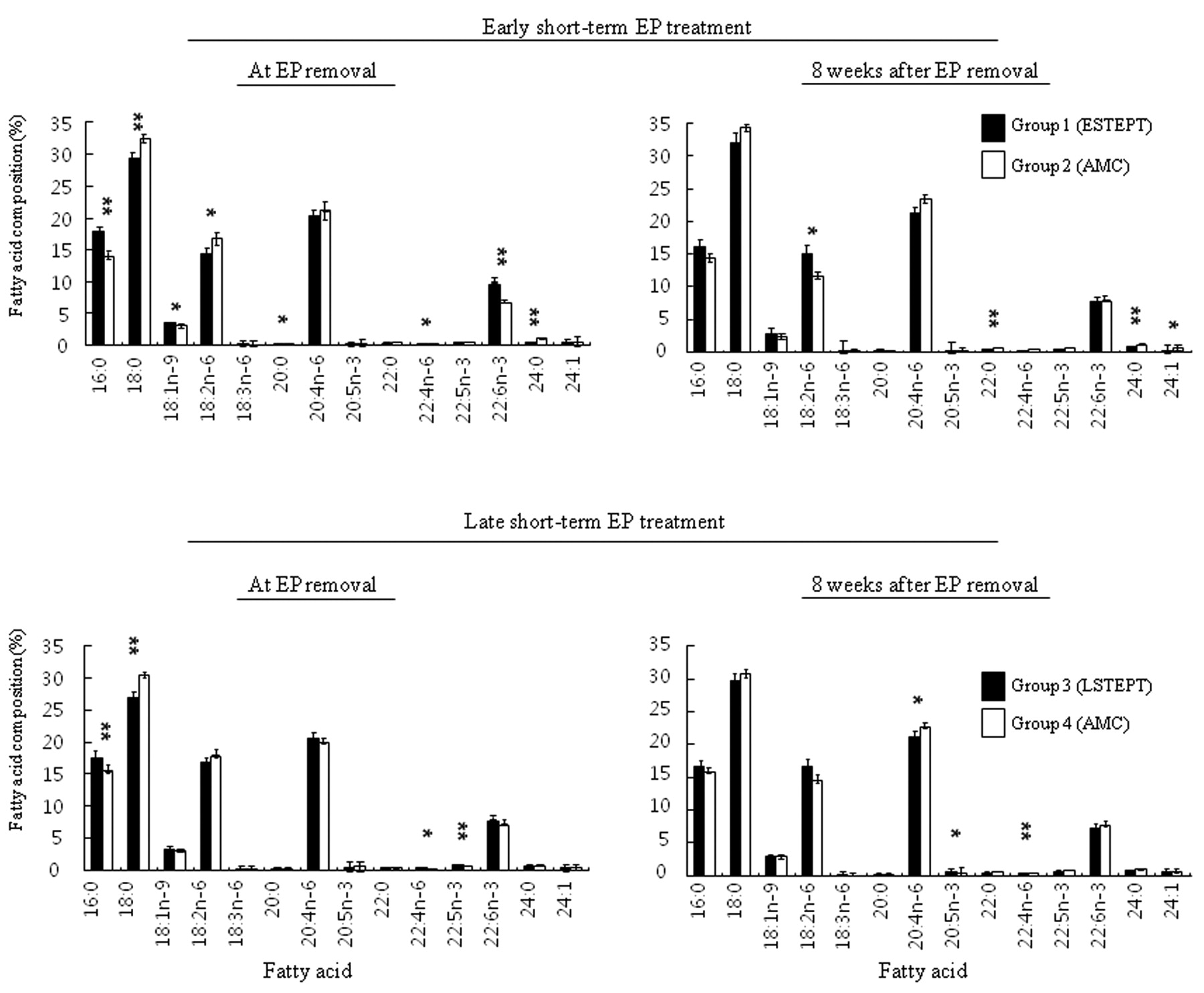
Fatty acid composition of serum phospholipids in MNU-treated rats exposed to ESTEPT or LSTEPT in comparison to respective age-matched controls. The individual fatty acid composition of serum phospholipids in the STEPT groups and their respective age-matched controls is presented in Figure 3. The prevalent fatty acids in serum phospholipids were palmitic acid (16:0), stearic acid (18:0), oleic acid (18:1n-9), linoleic acid (18:2n-6), arachidonic acid (20:4n-6), and DHA (22:6n-3). However, none of the changes in individual fatty acid composition (except DHA) were correlated with mammary carcinogenesis; there was a significant increase in DHA in the ESTEPT group at the time of EP removal (but not 8 weeks afterward). Next, fatty acids were expressed as SFA, MUFA, PUFA, n-6 PUFA, n-3 PUFA, and n-6/n-3 PUFA. When the levels of SFA, MUFA, and PUFA of the ESTEPT and LSTEPT groups were compared with the corresponding levels in respective age-matched controls, no significant difference was found in relation to mammary carcinogenesis (Tables II and III). However, a significant decrease in the n-6/n-3 PUFA ratio was seen in the ESTEPT group at the time of EP removal (but not 8 weeks afterward); the decrease was mainly due to a significant decrease in n-6 PUFA and a significant increase in n-3 PUFA (particularly DHA). However, in the LSTEPT group at the time of EP removal, no such significant change in the n-6/n-3 ratio was seen; a significant increase in n-3 PUFA did not result in a significant decrease in the n-6/n-3 ratio. Therefore, a transient drop in the n-6/n-3 ratio correlated with the suppression of mammary carcinogenesis in the ESTEPT group, but not in the LSTEPT group.
Discussion
This study demonstrated that ESTEPT (but not LSTEPT) effectively suppressed mammary carcinogenesis in female Lewis rats, which is consistent with previous reports (10, 11, 27). In rats, the mammary cancer-suppressing potential was weakened when the age of the rat at the first pregnancy was delayed (9), and this suppression was lost when STEPT was applied to middle-aged rats (24-27 weeks of age) (11, 27). Although the time of the initiation of carcinogenic insults in human breast cancer patients is unknown, the breast tissue of young women is more sensitive to radiation-induced carcinogenesis than that of older women (28); this observation suggests that carcinogenic initiation to human breast epithelial cells most likely occurs early in life. In rats, carcinogenic (MNU) initiation at puberty or adolescence (4 or 7 weeks of age) and STEPT at middle age (24 to 27 weeks of age) accelerated the occurrence of small-sized tumors and resulted in a higher incidence of palpable mammary tumors compared to age-matched controls (11, 27). Carcinogen exposure at a young age and LSTEPT (long interval between carcinogenic initiation to STEPT; ≥17 weeks) may accumulate transformed cells in the mammary gland which may be more likely to contain transformed malignant cells.
Changes in fatty acid composition after early short-term oestrogen and progesterone treatment in MNU-treated female Lewis rats.
Changes in fatty acid composition after late short-term oestrogen and progesterone treatment in MNU-treated female Lewis rats.

Comparison of fatty acid composition in serum phospholipids of MNU-exposed rats with or without short-term oestrogen and progesterone treatment. Fatty acids are shown as the number of carbons:the number of double bonds. The first position of the double bond numbered from the methyl terminus is designated as n-3, n-6, or n-9. EP, Oestrogen and progesterone pellet; ESTEPT, early short-term oestrogen and progesterone treatment; AMC, age-matched controls; LSTEPT, late short-term oestrogen and progesterone treatment. *p<0.05 and **p<0.01, compared with respective AMCs.
In contrast, the interval of time between carcinogen exposure and ESTEPT is usually short (approximately 2 weeks) in animal experiments in which STEPT is followed by carcinogenic initiation (9). To compare the effect of age on the susceptibility of parity-mediated protection, the time interval between carcinogenic initiation to STEPT should be identical. In the present study, the interval between carcinogenic initiation to ESTEPT and LSTEPT was thus adjusted to 2 weeks to clearly evaluate the age effect on STEPT. A single injection of the mammotropic chemical carcinogen MNU can effectively induce mammary cancer in susceptible strains of rats; female prepubertal rats are more susceptible than adolescent rats (the time when mammotropic carcinogen is usually applied), while middle-aged (6-8 months of age) rats are rather resistant (29, 30). Here, the regular 50 mg/kg dose of MNU was given to 4-week-old prepubertal rats and a 50 mg/kg dose twice in a one-week interval to 22- and 23-week-old middle-aged rats. Despite the doubled MNU dose, a long latency was needed to obtain sufficient numbers of mammary tumors in MNU-treated, middle-aged rats. However, the present study clearly reproduced the human situation in that early full-term pregnancy (i.e. ESTEPT here) lowered the risk of mammary carcinogenesis, while the protective effect was lost when STEPT was delayed.
The study examined whether the fatty acid composition of serum phospholipid levels was related to mammary cancer risk. The changes in fatty acid composition in serum phospholipids can be related to changes in the dietary intake of fatty acids. However, all groups of rats in the present study (ESTEPT, LSTEPT, and age-matched controls) ingested the same rodent chow. Thus, changes in fatty acid composition are not diet-related. In a nested case–control study, no significant associations were found between the composition of individual fatty acids, levels of SFA, MUFA, PUFA and the risk of breast cancer (22). Consistent with this previous study, no significant association was found between the serum phospholipid levels of SFA, MUFA, and PUFA and mammary cancer risk. There appears to be two types of fatty acids with opposing effects on mammary carcinogenesis: cancer-promoting n-6 PUFA and cancer-suppressing n-3 PUFA. Although there has been no association between serum phospholipid levels of n-3 or n-6 PUFA and human breast cancer risk (22, 31), there was a significant increase in n-3 PUFA levels in the ESTEPT and LSTEPT groups at the time of EP pellet removal (but not 8 weeks later), compared with respective age-matched controls.
The alteration of the n-6/n-3 ratio, rather than the amount of n-3 PUFA, seems to be important (32). The n-6/n-3 ratio consumed in the current Western diet is between 10:1 and 20:1; however, our ancestors consumed a diet with a ratio that was closer to 1:1. In a study of MNU-induced mammary carcinogenesis, a 1:1 n-6/n-3 diet significantly reduced the mammary cancer incidence and multiplicity, compared to diets with a 5:1 or higher n-6/n-3 PUFA ratio (33). In the present study, there was a transient but significant association between a decreased n-6/n-3 ratio, which was caused by ESTEPT but not by LSTEPT, and a decreased mammary cancer risk. During human pregnancy, the n-6/n-3 PUFA ratio in the plasma phospholipids decreases significantly (24). The maternal daily fat intake appears to be consistent during pregnancy (34); therefore, a decrease in the n-6/n-3 PUFA ratio cannot be explained by changes in the intake of dietary fatty acid. Rather, this decrease may be a maternal adaptation to the requirements of highly proliferating foetal tissue for development and growth, and it may provide self-protection against breast cancer. However, it was noteworthy that LSTEPT did not decrease the n-6/n-3 ratio or lower the mammary cancer risk.
Similar to changes in serum fatty acid composition, a significant increase in the concentration of n-3 PUFA and a decrease in the n-6/n-3 PUFA ratio occurs in the pregnant mouse mammary gland; mammary gland differentiation and β-casein expression is mediated through the Jak2/Stat5 signaling pathway (35). The organs and tissues of fat-1 transgenic mice capable of converting n-6 PUFA to n-3 PUFA contain abundant n-3 PUFA and reduced n-6 PUFA levels (a decreased n-6/n-3 PUFA ratio), and the mammary gland shows differentiation that is similar to that in early pregnancy (35). Thus, an increased n-3 PUFA level and a decreased n-6/n-3 PUFA ratio may induce a differentiation phenotype. In a human case-control study, decreased n-3 PUFA levels and increased n-6/n-3 PUFA ratios were seen in breast fat tissues from breast cancer patients compared with controls (36, 37). In rats, the fatty acid composition of serum reflects the fatty acid composition of the mammary gland (21). In addition to diet, early pregnancy (ESTEPT) may physiologically increase the level of n-3 PUFA and decrease the n-6/n-3 ratio; these changes in the fatty acid composition of the blood and mammary tissue may play a crucial role in mammary carcinogenesis.
With regard to the fatty acid composition of plasma cholesteryl esters, men have significantly lower DHA concentrations than women, and women taking oral contraceptives tend to have higher DHA concentrations than women not taking them (38). Administration of oral ethinyl estradiol increases DHA levels by 43%; oestrogens cause higher DHA concentrations probably by up-regulating DHA synthesis from β-linolenic acid (18:3 n-3) (38). In the present study, a similar effect of the EP pellet was found on DHA in young rats but not in old rats. If the up-regulation of DHA synthesis is the only reason for the increased DHA levels found in the ESTEPT group, it is unclear why DHA concentrations did not increase in the LSTEPT group. It is possible that the origin of increased DHA levels caused by oestrogen treatment was not dietary β-linolenic acid but rather the brain. Brain DHA content of postpartum dams fed a DHA-deficient diet was significantly decreased by 21% compared with that of age-matched virgin animals. Comparing rats fed a control diet, the decrement rates were reduced to 11% (one half of the decrease that occurs with a DHA-deficient diet; a nonsignificant change) (39). Interestingly, the brains of aged rats have significantly lower DHA concentrations in the phosphatidylethanolamine fraction than those of younger controls (40). Consequently, decreased brain levels of DHA in the LSTEPT group may be the reason why STEPT was unable to increase the blood levels of DHA; that is, there may be no available DHA in the brains of older rats.
The present study demonstrated a significantly lower n-6/n-3 ratio in serum phospholipids of rats exposed to ESTEPT (but not LSTEPT) compared to their respective age-matched controls. Although other mechanisms may be responsible for the effects of ESTEPT suppressing mammary carcinogenesis, early pregnancy (ESTEPT)-mediated mammary cancer prevention was correlated with a decreased n-6/n-3 ratio and an increased level of DHA. Further studies are needed to determine whether a change in the serum fatty acid composition is the primary event that controls mammary carcinogenesis after STEPT.
Acknowledgements
The Authors thank Ms. T. Akamatsu, H. Hamatani and S. Takebe for their technical help and Ms. A. Shudo for manuscript preparation. This study was supported in part by a Grant-in-Aid for Scientific Research (C) from the Japan Society for the Promotion of Science (JSPS) (21591683), a grant from the Osaka Community Foundation, Fujii Memorial Osaka Foundation for Promotion of Fundamental Medical Research, and a grant from Sonoko Morimoto Foundation of the Kansai Medical University Alumni Association.
- Received April 12, 2010.
- Revision received May 26, 2010.
- Accepted May 28, 2010.
- Copyright © 2010 International Institute of Anticancer Research (Dr. John G. Delinassios), All rights reserved




{kind=link}
{kind=link}
{kind=link}